

Effects of Seedling Tray Bagging Duration on Growth and Quality of In-forest Cultivated Anoectochilus roxburghii
-
摘要:目的 明确林下穴盘袋栽对金线莲生长和品质的影响,筛选出适宜金线莲生长的套袋时长,为金线莲科学种植、品质提升提供理论依据。方法 以南靖大叶金线莲为试验材料,在南靖县葛山村试验地设置7个不同套袋时长处理,以落地栽培为对照(CK),研究穴盘袋栽及不同套袋时长对金线莲生长及品质的影响。结果 (1)穴盘袋栽金线莲可以提高保存率、株高、叶片数、叶长、叶宽、茎节数、每节长、根数和根长,与对照相比,穴盘袋栽套袋150 d处理的保存率增加26.42%,株高增加26.47%,叶片数增加36.36%,每节长增加18.41%,平均单株鲜重增加8.32%,且存在显著差异;(2)穴盘栽培较落地栽培可显著提升金线莲可溶性糖含量,套袋时间过长不利于金线莲叶绿素合成,而对可溶性蛋白含量无显著影响;(3)金线莲穴盘栽培较落地栽培有利于总多糖、总黄酮、总生物碱和总氨基酸含量的积累与合成,分别增加11.85%、5.67%、3.70%、17.50%。但套袋时间过长则不利于金线莲总生物碱、金线莲苷、总酚含量提升,套袋150 d与套袋0 d处理的金线莲总黄酮、总生物碱、总氨基酸、总多糖含量达到的最高值之间无显著差异。结论 穴盘袋栽较落地栽培有利于金线莲生长和品质提升,过长的袋栽时长不利于金线莲活性成分的合成,综合考虑,穴盘袋栽以套袋时长150 d较为适宜,金线莲生长和品质较佳。Abstract:Objective Effects of duration of the seedling tray bagging on the growth and quality of in-forest cultivated Anoectochilus roxburghi were analyzed.Method Under tree shade in the forest, Nanjing large leaf A. roxburghii seedlings were cultivated with the conventional ground planting (CK) or utilization of the seedling tray bagging method for 7 different lengths of time at an experimental site in Geshan Village, Nanjing County. Effects of the bagging and bagging duration on the growth and quality of the plants were evaluated.Result (1) The seedling tray bagging significantly increased the survival rate, plant height, leaf number, leaf length, leaf width, stem node number, node length, root number, and root length of the A. roxburghii plants. Compared with CK, 150 d bagging afforded the plants 26.42% higher on survival rate, 26.47% taller on height, 36.36% more on number of leaves, 18.41% longer on node, and 8.32% on average plant fresh weight. (2) The bagging significantly raised the soluble sugar content in A. roxburghii over direct ground planting. However, too long a bagging duration was detrimental to chlorophyll synthesis of the plant, even though no significant effect on protein content. (3) The bagging facilitated the plant accumulation and synthesis of polysaccharides, flavonoids, alkaloids, and amino acids, as shown by 11.85%, 5.67%, 3.70%, and 17.50% increases, respectively, over the conventional ground cultivation. However, being held in the bags for a prolonged period, the plants ceased to rise on the contents of total alkaloids, kinsenoside, and total phenols. The peak contents of flavonoids, alkaloids, amino acids, and polysaccharides did not significantly differ on the plants bagged for 150 d or 0 d either.Conclusion The seedling tray bagging practice benefitted the growth and quality of A. roxburghii. However, prolonged bagging was not conducive to the synthesis of functional ingredients in the plant. It appeared that bagging for 150 d was optimal for the cultivation of high quality A. roxburghii in the woods.
-
0. 引言
【研究意义】高产是水稻选育的首要目标,研究表明,水稻的产量是由每穗粒数、千粒重等多个产量因子构成的复杂农艺性状,同时水稻的农艺性状多为数量性状,各性状间相互影响,导致难以量化不同农艺性状对单株产量的贡献大小[1-2]。此外,不同的杂交组合、气候和栽培条件下,其产量构成因子对产量的贡献以及相关程度也不同[3-5]。因此,结合地区实际情况,探明不同农艺性状和产量的关系,可为高产杂交水稻品种选育提供理论依据。水稻种质资源是育种的物质基础,水稻育种工作对种质资源的利用最终归结为对性状配合力的选择[6-7]。杂交亲本配合力的高低直接影响杂交品种的产量,利用水稻杂种优势的育种实质上是配合力育种[8]。因此,了解和掌握水稻三系不育系主要农艺性状的配合力,能够有效评价不育系的育种潜力,对强优势杂交水稻组合选配具有重要意义。【前人研究进展】水稻配合力指一般配合力和特殊配合力,两者在当今水稻杂交育种是首要考虑的因素。一般配合力与杂交组合的竞争优势存在着显著正相关,且特殊配合力大小能够充分反映杂种优势的大小[9-10]。一般配合力高的亲本基础上需再组配特殊配合力高的亲本才能获得优良杂交品种[11-12]。但一般配合力低的双亲同样能配出特殊配合力高的组合[7]。由此可见,亲本的配合力在水稻杂交育种中起到决定性作用,如何利用亲本的一般配合力和特殊配合力是利用杂种优势的关键。研究表明,农艺性状对水稻产量影响很大,有效穗数、穗长、每穗粒数、千粒重是影响水稻产量的重要因素[13-14]。结实率是影响大穗杂交稻产量的主要因素[15]。总粒数对早籼产量影响最大,总粒数越多产量越高[16]。综上所述,由于品种、栽培方式、地理环境等因素的差异,水稻主要农艺性状与产量的遗传关系表现出多样性。【本研究切入点】前人研究均是基于特定区域应用特定的品种,对各自区域的育种以及生产实践具有一定的指导意义。因此,掌握川南中籼稻区不育系配合力及遗传力的规律,有助于正确选择亲本,组配强优品种。此外,川南水稻农艺性状与产量间关系的研究鲜有报道,明确影响水稻产量的关键农艺性状,有助于指导本地区的亲本改良。【拟解决的关键问题】通过对6个不育系的主要农艺性状进行配合力分析及遗传率估算,评价新不育系的育种潜力,为新不育系的利用和强优组合的组配提供依据;同时,通过对不同水稻组合的多个农艺性状与产量的相关及通径分析,探讨各性状和单株产量间的关系,以明确影响单株产量的关键农艺性状。
1. 材料与方法
1.1 试验材料
供试的6个不育系:宜香1A(1A,本单位选育)、神9A(9A,重庆市农业科学院引进),4个新选不育系分别是五良A(A1)、酒都A(A2)、Z1A(A3)和1281A(A4);供试的6个恢复系:蜀恢527(R527,四川农业大学引进)、成恢727(R727,四川省农业科学院引进)、雅恢2115(R2115,四川农业大学引进)、绵恢6139(R6139,绵阳市农业科学研究院引进)、中种恢2877(R2877,中国种子集团有限公司引进)、宜恢3003(R3003,本单位选育)。
1.2 试验方法
1.2.1 杂交组合制备与种植
2020年夏,在宜宾市农业科学院大观镇试验基地,按照NC II不完全双列杂交组配组合36个;2021年5月28日对36个试验组合进行种植。采用随机区组设计,3次重复,行株距33.3 cm×16.7 cm,每行栽8株,每份材料栽3行,单株栽插。田间农事管理同当地大田生产,施肥水平较低。
1.2.2 性状调查
水稻成熟后,每个小区随机选取5株测量株高(Plant height,PH),随机选取5株风干考种,调查穗长(Panicle length,PL)、千粒重(1000-grain weight,TGW)、每穗实粒数(Filled Spikelets per panicle,FSP)、每穗颖花数(Spikelets per panicle,SP)、结实率(Seed setting rate,SSR)、单株有效穗数(Tillers per plant,TP)和单株产量(Grain weight per plant,GWP)等农艺性状。
1.3 统计分析
利用Excel 2013软件和DPS数据分析软件进行各农艺性状的方差分析和配合力分析、相关性分析及通径分析[17]。
2. 结果与分析
2.1 主要农艺性状的配合力方差分析
表1列出了各个性状的配合力方差(F值)。从表1可知,各性状在组合间的方差均达到极显著水平,说明各个性状在组合间存在明显的遗传差异,可以进行配合力方差分析。组合间的遗传差异来源于不育系、恢复系以及不育系和恢复系的互作效应。参试不育系除每穗颖花数外,其余7个性状一般配合力方差均达到显著或极显著水平;恢复系除单株产量外,其余7个性状一般配合力方差均达到显著或极显著水平;除株高外其余7个性状的特殊配合力均达到显著或极显著差异水平,以上结果说明基因加性效应和基因互作的非加性效应对研究的8个农艺性状有着重要影响,具体可以进行一般配合力(GCA)和特殊配合力(SCA)的效应分析。
表 1 主要农艺性状的配合力方差分析Table 1. Analysis of variance on combining ability of main agronomic traits变异来源
Source ofvariatioin自由度
df株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP单株产量
GWP组合 Combination 35 18.1665** 22.4393** 398.3065** 10.3568** 13.9383** 4.5617** 5.4225** 3.8326** 恢复系 R 5 18.5998** 4.015** 82.6418** 9.7716** 12.6394** 4.9701** 3.01* 2.4425 不育系 S 5 58.4764** 19.0802** 31.5235** 2.7176* 1.6981 7.6693** 5.2619** 3.9612** 恢复系×不育系 R×S 25 1.5494 5.5908** 23.3973** 4.1453** 5.0455** 1.8103* 2.86** 2.3526** “*”和 “**”分别表示显著差异(P<0.05)和极显著差异(P<0.01)。df:自由度 。
* and ** represent significant difference (P <0.05) and extremely significant difference (P <0.01), respectively. df: degrees of freedom.2.2 不育系主要农艺性状一般配合力效应分析
由表2可知,不同不育系同一性状或相同不育系不同性状的GCA效应值存在明显差异,表明6个不育系在各性状上的加性效应大小不同,不育系间的同一性状及同一性状在不同不育系间存在明显的遗传差异。在参试的6个不育系中,株高、穗长、千粒重、每穗实粒数、每穗颖花数、结实率、单株有效穗、单株产量GCA效应值从大到小分别为A3(4.4103)、1A(4.2823)、1A(7.457)、9A(4.0523)、A3(2.6181)、9A(2.4747)、A1(7.6541)、1A(4.9982);4个新不育系中, A1除千粒重的GCA呈现负效应外,其余性状的GCA均呈现为正效应; A2在株高、穗长、千粒重、单株有效穗数、单株产量等性状的GCA呈现为正效应;A3在株高、穗长、每穗实粒数、每穗颖花数等性状的GCA呈现为正效应;A4在多个农艺性状上的GCA均呈现负效应。
表 2 不育系农艺性状一般配合力效应值Table 2. GCA of agronomic traits in male sterile rice不育系
S株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP单株产量
GWP1A 3.4508 4.2823 7.457 −2.0734 −1.6291 −0.4184 −0.3976 4.9982 9A −3.0294 −8.9093 −3.4697 4.0523 1.5552 2.4747 −5.169 −4.6277 A1 0.6664 3.1679 −4.9266 1.7651 1.2399 0.5676 7.6541 4.5002 A2 0.4749 1.1578 5.425 −4.2365 −3.7746 −0.5896 2.4354 3.3596 A3 4.4103 1.8677 −1.8978 2.41 2.6181 −0.1243 −3.5288 −2.7706 A4 −5.9731 −1.5664 −2.5879 −1.9176 −0.0096 −1.9099 −0.994 −5.4598 2.3 杂交组合特殊配合力效应分析
由表3可知,36个组合中,株高SCA效应值为负值的组合有17个,负效应较高的组合是A3×R3003和A2×R727,效应值均在−1.7以下;穗长SCA效应值为正值的有19个,较高的是A2×R6139、9A×R727和A1×R727,效应值均在3.7以上;千粒重SCA效应值为正值的有18个,较高的是A1×R2115、9A×R2877,效应值均在3.8以上;每穗实粒数SCA效应值为正值的有23个,较高的是A2×R2877、9A×R3003,效应值均在6.3以上;每穗颖花数SCA效应值为正值的有20个,较高的为9A×R3003和1A×R2877,效应值均在5.19以上;结实率SCA效应值为正值的有18个,较高的是A2×R2877、A3×R527,效应值均在1.5以上;单株有效穗数SCA效应值为正值的有18个,较高的是9A×R3003、A1×R727,效应值均在6.9以上;单株产量SCA效应值为正值的有17个,较高的是1A×R2115、A1×R727,效应值均在5.8以上。
表 3 杂交组合主要农艺性状的特殊配合力效应值Table 3. SCA of agronomic traits in hybrids组合
Combination株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP单株产量
GWP1A×R527 −0.4269 1.4426 −0.0192 1.9138 3.2788 −1.383 1.8887 3.9176 1A×R727 1.6077 −2.4621 1.3994 −4.9336 −5.6306 0.7786 3.3797 −0.3813 1A×R2115 0.918 1.2032 −1.5144 5.4748 4.6071 0.8519 2.9324 7.4165 1A×R6139 −0.5877 −0.8812 1.0543 0.023 −1.0591 1.1414 −1.839 −0.7411 1A×R2877 −1.5482 −1.0711 −3.3547 4.568 5.1956 −0.512 −3.9264 −2.3636 1A×R3003 0.0372 1.7687 2.4345 −7.046 −6.3918 −0.8768 −2.4354 −7.8481 9A×R527 0.4865 0.4685 1.016 0.0192 −0.9609 1.0925 −4.9702 −4.2303 9A×R727 −0.5546 3.7457 −1.4761 1.3816 2.5558 −1.2264 −7.0577 −7.4304 9A×R2115 0.3885 −1.133 −1.6294 2.3229 3.4048 −1.1412 −5.7157 −5.6665 9A×R6139 −0.6333 −1.1124 −0.901 −1.4146 −1.0669 −0.3164 4.7217 2.5039 9A×R2877 0.2292 −2.0452 3.8914 −8.6839 −9.2064 0.2585 4.4235 0.3224 9A×R3003 0.0838 0.0764 −0.901 6.3747 5.2736 1.333 8.5984 14.501 A1×R527 −1.4814 −4.1791 −2.8179 4.3785 3.2229 1.0409 1.8887 3.9722 A1×R727 −0.2186 3.7045 −1.8594 0.7344 0.1362 0.5618 6.9583 5.8807 A1×R2115 0.1486 2.8872 5.3483 −6.0131 −6.1936 0.2506 −1.5408 −2.0713 A1×R6139 −0.5853 −2.2433 −2.2045 2.0803 1.4112 0.5598 2.6342 1.8774 A1×R2877 0.4673 −0.3777 1.2077 −3.5822 −1.8479 −1.7042 −3.0318 −5.9717 A1×R3003 1.6696 0.2084 0.3259 2.4022 3.2713 −0.709 −6.9085 −3.6874 A2×R527 0.3401 0.357 1.3227 −3.5353 −1.9702 −1.5489 0.8449 −1.8464 A2×R727 −1.755 −4.241 −2.0895 1.0801 1.6282 −0.442 5.0199 4.6309 A2×R2115 1.2069 1.9503 1.2077 −1.5377 −1.9771 0.69 2.7833 2.3272 A2×R6139 −0.2038 3.9026 0.786 4.5353 4.1638 0.4997 −5.5666 −0.0086 A2×R2877 0.0857 0.5923 −2.0128 7.2398 4.4098 2.6686 −0.497 5.2124 A2×R3003 0.3261 −2.5611 0.786 −7.7822 −6.2545 −1.8674 −2.5845 −10.3156 A3×R527 −0.2345 0.1424 −0.0958 −1.5232 −3.0896 1.5742 0.5467 −1.0367 A3×R727 −0.6996 −0.5675 1.7828 0.0502 0.3402 −0.368 0.2485 2.0091 A3×R2115 −1.0984 −0.8647 −1.131 2.7596 2.7697 −0.1519 1.5905 3.0212 A3×R6139 1.6206 −2.9739 0.2875 −6.0238 −5.315 −0.8061 1.2922 −4.2689 A3×R2877 2.1981 1.5912 −0.2109 0.1322 1.8264 −1.496 −1.6899 −2.4022 A3×R3003 −1.7862 2.6726 −0.6326 4.6049 3.4683 1.2478 −1.9881 2.6774 A4×R527 1.3163 1.7687 0.5943 −1.253 −0.481 −0.7757 −0.1988 −0.7765 A4×R727 1.6202 −0.1795 2.2428 1.6873 0.9702 0.696 −8.5487 −4.7091 A4×R2115 −1.5635 −4.0429 −2.2812 −3.0066 −2.6109 −0.4995 −0.0497 −5.0271 A4×R6139 0.3895 3.3082 0.9776 0.7998 1.8661 −1.0784 −1.2425 0.6372 A4×R2877 −1.432 1.3105 0.4792 0.3259 −0.3776 0.7852 4.7217 5.2028 A4×R3003 −0.3305 −2.1649 −2.0128 1.4464 0.6332 0.8724 5.3181 4.6727 2.4 双亲在农艺性状上的方差贡献率及遗传力比较分析
为研究不育系、恢复系及其互作对F1代8个性状的影响,根据随机模型估算各个性状的基因型方差遗传分量、一般配合力和特殊配合力基因型遗传方差,以及一般配合力和特殊配合力基因型遗传方差在总遗传方差中的比重,一般配合力遗传方差中不育系、恢复系所占比重。从表4可知,8个农艺性状的VG均大于50%,其中株高、千粒重的VG达到了95%以上,表明双亲的加性效应对F1的8个农艺性状起主导作用。在一般配合力基因型遗传方差中,不育系和恢复系所占比重因性状而异,在株高、穗长、单株有效穗、结实率和单株产量等5个性状上不育系占比更高,说明不育系对F1相应性状起主导作用;而在千粒重、每穗实粒数和每穗颖花数3个性状上恢复系占比更高,说明恢复系对相应性状起主导作用。
表 4 主要农艺性状的基因型方差、父母本及其基因互作对F1各性状方差的贡献率和遗传力Table 4. Contributions of genotypic variance of agronomic traits, parents, and gene interactions to trait variance and heritability of F1 hybrids项目
Project株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP单株产量
GWP基因型方差 Genotypic variance 恢复系 R lines 6.2001 0.2634 5.7272 85.9889 139.6384 0.778 0.0445 1.0054 不育系 S lines 20.248 1.5798 2.1412 16.8379 8.3752 1.3069 0.0943 2.0639 互作 R×S 0.7495 0.4305 0.4029 44.6293 57.7158 0.5263 0.0863 2.4042 贡献率 Contribution VG/% 97.24 81.07 95.13 69.73 71.95 79.85 61.65 56.07 Vg1/% 22.79 11.59 69.24 58.31 67.88 29.8 19.76 18.37 Vg2/% 74.45 69.48 25.89 11.42 4.07 50.05 41.89 37.7 Vs/% 2.76 18.93 4.87 30.27 28.05 20.15 38.35 43.93 遗传力 Heritability 广义遗传率(hB2)/% 86.92 88.99 99.35 77.6 82.78 57.27 61.78 50.65 狭义遗传率(hN2)/% 84.52 72.14 94.51 54.11 59.56 45.72 38.09 28.4 VG为双亲一般配合力基因型方差占总方差的比重,Vg1为恢复系一般配合力基因型方差占总方差的比重,Vg2为不育系一般配合力基因型方差占总方差的比重,Vs为特殊配合力基因型方差占总方差的比重。
VG: Proportion of genotype variance of GCA in total variance; Vg1: genotype variance of GCA of restorer lines in total variance; Vg2: genotype variance of GCA of male sterile lines in total variance; Vs: proportion of genotype variance of SCA in total variance.遗传力是反映该性状的遗传变异占总变异的百分比。从表4可知,株高、穗长、千粒重和每穗颖花数等4个性状的广义遗传力均大于80%,表明这些性状主要受到遗传的影响,其中千粒重、株高、穗长的狭义遗传率也较高,均大于70%,说明三者遗传主要受到基因加性效应的影响,其性状易于稳定遗传给后代。结实率、单株有效穗数和单株产量的广义遗传率和狭义遗传率都相对较低,表明这些性状易受环境影响,在后代遗传中的稳定性较差。8个性状的狭义遗传率大小为:千粒重>株高>穗长>每穗颖花数>每穗实粒数>结实率>单株有效穗数>单株产量。
2.5 供试不育系测交组合的主要农艺性状平均表现
测交组合F1代的主要农艺性状平均表现是衡量亲本优秀与否的重要指标之一。从表5可知,株高较高的是A3、1A组配的组合,平均值分别达120.85 cm、119.74 cm;穗长较高的是1A和A1组配的组合,平均值分别达28.07 cm、27.77 cm;千粒重较高的是1A和A2组配的组合,平均值分别达31.14 g、30.55 g;每穗实粒数较高的是9A和A3组配的组合,平均值分别达167.72粒、165.07粒;每穗颖花数较高的是A3和9A组配的组合,平均值分别达196.74 粒、194.70粒;结实率较高的是9A和A1组配的组合,平均值分别达86.15%、84.55%;单株有效穗数较高的是A1和A2组配的组合,平均值达8.02个、7.63个;单株产量较高的是1A和A2组配的组合,平均值分别达36.31 g、36.14 g。在新不育系中, A1在株高、穗长、每穗实粒数、每穗颖花数、结实率、单株有效穗数、单株产量等性状上平均表现高于试验均值,千粒重低于试验均值;A2在株高、穗长、千粒重、单株有效穗数、单株产量等性状平均表现高于均值;A3在株高、穗长、每穗实粒数、每穗颖花数等性状平均表现高于均值。
表 5 供试不育系测交组合F1代主要农艺性状的平均表现Table 5. Average performance of agronomic traits in F1 hybrids of male sterile rice组合
Combination株高
PH/cm穗长
PL/cm千粒重
TGW/g每穗实粒数
FSP每穗颖花数
SP结实率
SSR/%单株有效穗数
TP单株产量
GWP/g1A/*** 119.74 28.07 31.14 157.85 188.60 83.72 7.42 36.31 9A/*** 112.24 24.52 27.98 167.72 194.70 86.15 7.07 32.98 A1/*** 116.52 27.77 27.55 164.03 194.10 84.55 8.02 36.14 A2/*** 116.30 27.23 30.55 154.36 184.48 83.57 7.63 35.74 A3/*** 120.85 27.42 28.43 165.07 196.74 83.96 7.19 33.62 A4/*** 108.83 26.50 28.23 158.10 191.70 82.46 7.38 32.69 均值Average 115.75 26.92 28.98 161.19 191.72 84.07 7.45 34.58 ***代表6个恢复系。
*** represents 6 restorer lines.2.6 主要农艺性状相关性及通径分析
相关系数是反映各农艺性状和产量间的线性关系,可以预测各农艺性状对产量的影响。从表6可见,单株产量与其他7个农艺性状的相关程度是千粒重>穗长>单株有效穗数>结实率>株高>每穗实粒数>每穗颖花数,其中千粒重、穗长、单株有效穗数和结实率与单株产量呈显著或极显著的正相关,株高和每穗实粒数与单株产量正相关但不显著,每穗颖花数与单株产量负相关且不显著。以上结果说明千粒重、穗长、单株有效穗和结实率对单株产量有着重要的影响,对任一性状的改良都会促进产量的提高。
表 6 组合农艺性状间的相关系数Table 6. Correlation coefficients between agronomic traits of hybrids农艺性状
Traits株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP单株产量
GWP株高 PH 1 穗长 PL 0.6243** 1 千粒重 TGW 0.47** 0.4346** 1 每穗实粒数 FSP −0.3671* −0.2986 −0.6192** 1 每穗颖花数 SP −0.3988* −0.2438 −0.7011** 0.963** 1 结实率 SSR 0.0714 −0.2154 0.2148 0.2781 0.0094 1 单株有效穗数 TP 0.0955 0.2235 −0.1924 −0.2365 −0.1906 −0.2029 1 单株产量 GWP 0.315 0.4493** 0.4733** 0.0179 −0.09 0.4007* 0.4111* 1 *和**分别表示显著差异(P<0.05)和极显著差异(P<0.01)。
* and ** represent significant difference (P<0.05) and extremely significant difference (P<0.01), respectively.为进一步明确本试验中7个农艺性状对单株产量的相对重要性,对相关系数进行了通径分析,结果见表7。除株高外其余性状对单株产量的直接通径系数为正值,表明这6个性状对单株产量都有直接的正效应,在保持其他条件不变下,通过提高6个农艺性状中任意一个性状,均能提高单株产量。7个农艺性状直接通径系数从大到小为千粒重>单株有效穗数>每穗实粒数>每穗颖花数>结实率>穗长>株高,其中千粒重、单株有效穗数对单株产量的直接作用达到极显著水平,其余性状对单产的直接作用均不显著,表明千粒重和单株有效穗数是影响单株产量的关键因素。根据间接通径系数结果,千粒重通过对穗长和结实率的正调控,对每穗实粒数、每穗颖花数和单株有效穗数的负调控来间接影响单株产量;单株有效穗数通过对穗长的正调控,对株高、千粒重、每穗颖花数和每穗实粒数的负调控来间接影响单株产量,表明农艺性状间存在相互促进与制约的关系,最终共同影响单株产量。
表 7 各性状与单株产量的直接和间接通径系数Table 7. Direct and indirect path coefficients on traits to yield per plant因素
Factors直接系数
Direct coefficient与产量的相关系数
Correlation coefficient
with yield间接系数 Indirect coefficient 株高
PH穗长
PL千粒重
TGW每穗实粒数
FSP每穗颖花数
SP结实率
SSR单株有效穗数
TP株高 PH −0.0055 0.315 0.0112 0.5826 −0.2712 −0.095 0.008 0.0849 穗长 PL 0.018 0.4493** −0.0034 0.5388 −0.2206 −0.0581 −0.024 0.1987 千粒重 TGW 1.2398** 0.4733** −0.0026 0.0078 −0.4575 −0.1671 0.0239 −0.1711 每穗实粒数 FSP 0.7388 0.0179 0.002 −0.0054 −0.7677 0.2295 0.031 −0.2103 每穗颖花数 SP 0.2383 −0.09 0.0022 −0.0044 −0.8692 0.7115 0.001 −0.1694 结实率 SSR 0.1114 0.4007* −0.0004 −0.0039 0.2663 0.2054 0.0022 −0.1803 单株有效穗数 TP 0.889** 0.4111* −0.0005 0.004 −0.2386 −0.1748 −0.0454 −0.0226 决定系数R2=0.994966,剩余通径系数=0.070952。
Coefficient of determination, R2=0.994966; residual path coefficient=0.070952.3. 讨论
3.1 杂交组合8个主要农艺性状的遗传特点
本研究中8个农艺性状在不育系或恢复系或两者间的一般配合力方差均达到显著或极显著水平,除株高外其他7个农艺性状的特殊配合力方差均达到显著或极显著水平,说明多数农艺性状是受到加性效应和非加性效应的共同影响。基因型方差及贡献率的估算结果表明,8个农艺性状均是基因的加性效应主导,且亲本的一般配合力与特殊配合力相互独立,无明显对应关系,这与前人研究[11,18-20]基本一致。遗传力估算结果表明株高、穗长、千粒重的遗传力较高,说明三者受到环境的影响较小,容易通过亲本的选择获得期望的后代,这与刘金波[7]、潘清洁[11]等研究结果相似。
刘龙钦等研究认为有效穗、结实率、千粒重受不育系影响较大[21]。李双等研究认为有效穗数、千粒重、单株产量、株高和穗长受不育系影响较大[22]。陈锋等研究认为株高、穗长和千粒重受不育系影响较大,穗长、每穗粒数和千粒重受恢复系影响更大[23]。而本研究结果表明,在株高、穗长、单株有效穗、结实率和单株产量等性状主要受到不育系的影响,而千粒重、每穗实粒数和每穗颖花数主要受到恢复系的影响。可见,农艺性状遗传具有复杂性、多样性的特点,不同学者针对不同材料研究会得出不同的结论,因此在实际育种工作中应考虑到所用材料的遗传特点,不能一概而论。
3.2 水稻农艺性状和单株产量的关系
水稻的产量形成是个系统工程,而影响水稻产量的各个性状间又存在复杂的相关关系,对任何一个性状进行变动,都将不同程度地影响其他性状[3,24]。本研究中的相关分析结果表明,千粒重、穗长、单株有效穗数和结实率与单株产量均呈显著或极显著正相关,说明四者对单株产量有着重要的影响,优化任一性状均能明显提升产量,这与育种经验相吻合。通径分析结果表明,千粒重、单株有效穗数对单株产量的直接作用达到极显著水平,这与相关分析结果不完全相同,但结合两者我们不难发现千粒重和单株有效穗数是影响单株产量的关键因素,这与杨久[25],李雪梅[26]等的研究结果相似。通径分析结果还揭示了水稻农艺性状间的复杂关系,水稻农艺性状间通过相互促进与制约共同影响单株产量。因此,对关键农艺性状进行选择时,应考虑各农艺性状之间的关联性,兼顾其他性状,如在本地区进行高产育种时,在注重千粒重和单株有效穗数选择的同时,也应兼顾穗长、结实率等农艺性状考察。
3.3 不育系表现及评价
从几个新不育系的一般配合力来看,五良A多个性状的一般配合力均较好,易组配出分蘖能力强、结实率高、穗粒数多、产量高的组合,但其千粒重的一般配合力较差,组配时应选大粒父本进行互补;酒都A千粒重、结实率、单株有效穗数和单株产量的一般配合力较好,但每穗颖花数、每穗实粒数和结实率的一般配合力表现一般;Z1A穗长、每穗实粒数和每穗颖花数的一般配合力较好;1281A株高具有优势,有待进一步发掘利用。从特殊配合力来看,五良A×成恢727组合多个性状的特殊配合力均较好,综合表现最好,其次是酒都A×中种恢2877、1281A×中种恢2877。综上可知,未发现不育系8个农艺性状的一般配合力均表现优秀,因此,育种时应根据育种目标进行合理的亲本组配从而获得优良组合。
4. 结论
不育系五良A和酒都A的一般配合力和特殊配合力都较高,两者分别在单株有效穗数、千粒重方面具有较强优势,在本地区具有良好的应用前景。川南地区生态条件区域间差异大、垂直分布明显、农业立体性强、旱涝灾害频繁,加之光能不足,阴雨和雾日多,因此对培育高产作物组合提出了更高的要求[27]。在川南中籼稻区进行高产育种时,应重点考察千粒重和单株有效穗数两个性状,同时兼顾每穗实粒数、每穗颖花数、穗长等性状进行选育。在大田生产实践中,合理使用提高千粒重、有效穗数的栽培方法,将有效提高水稻产量。
-
![]()
图 1 金线莲穴盘袋栽
注:A,穴盘;B,穴盘+骨架;C,穴盘袋栽;D,穴盘袋栽管理。
Figure 1. Schematic diagram of seedling tray bagging for A. roburghii cultivation
Note: A, Seedling tray; B, Seedling tray and framework; C, Seedling tray bagging cultivation; D, Seedling tray bagging cultivation management.
![]()
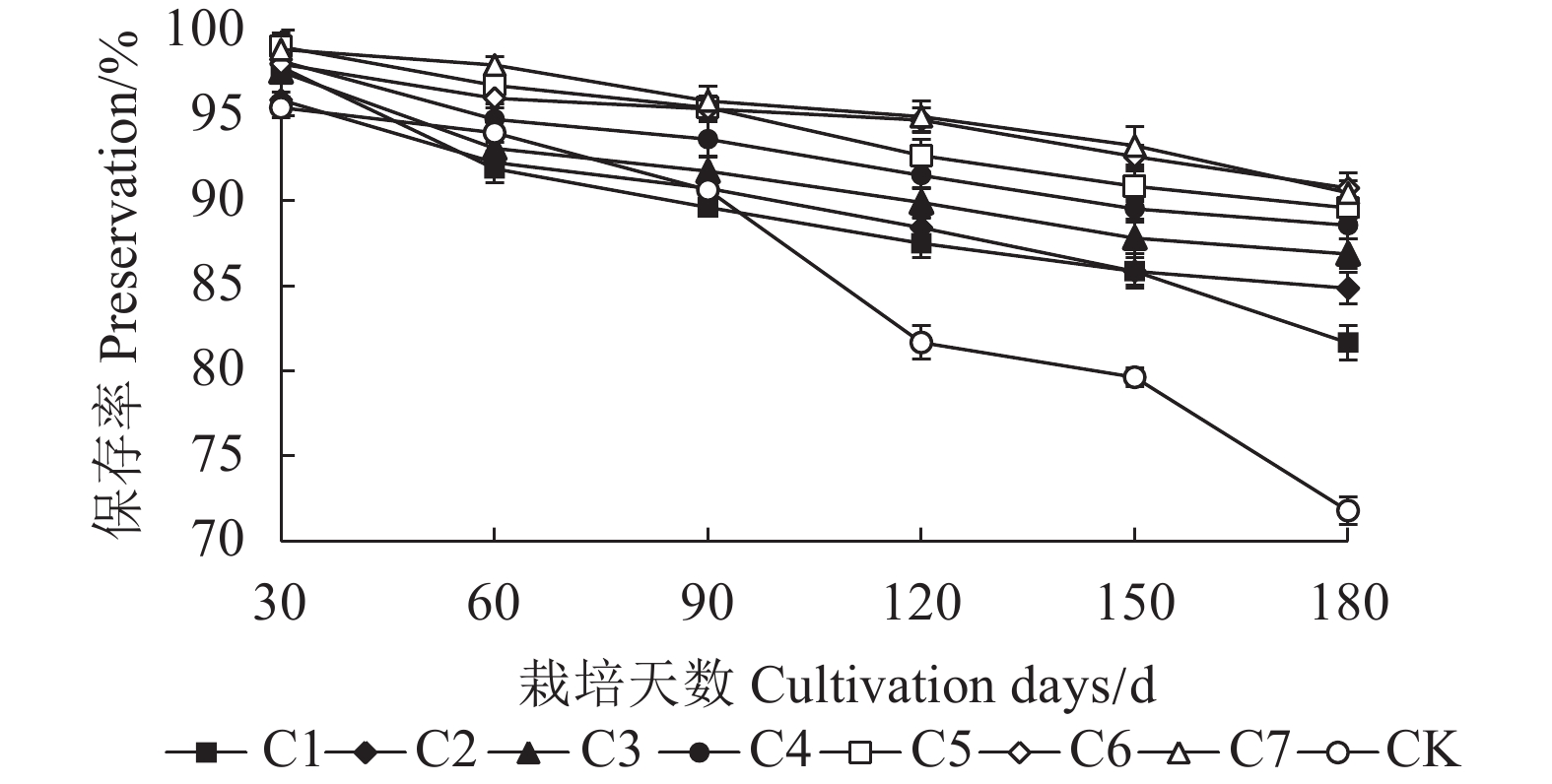
图 2 不同穴盘套袋时长金线莲保存率
Figure 2. Survival rate of A. roburghii for different duration of Seedling tray bagging cultivation
![]()
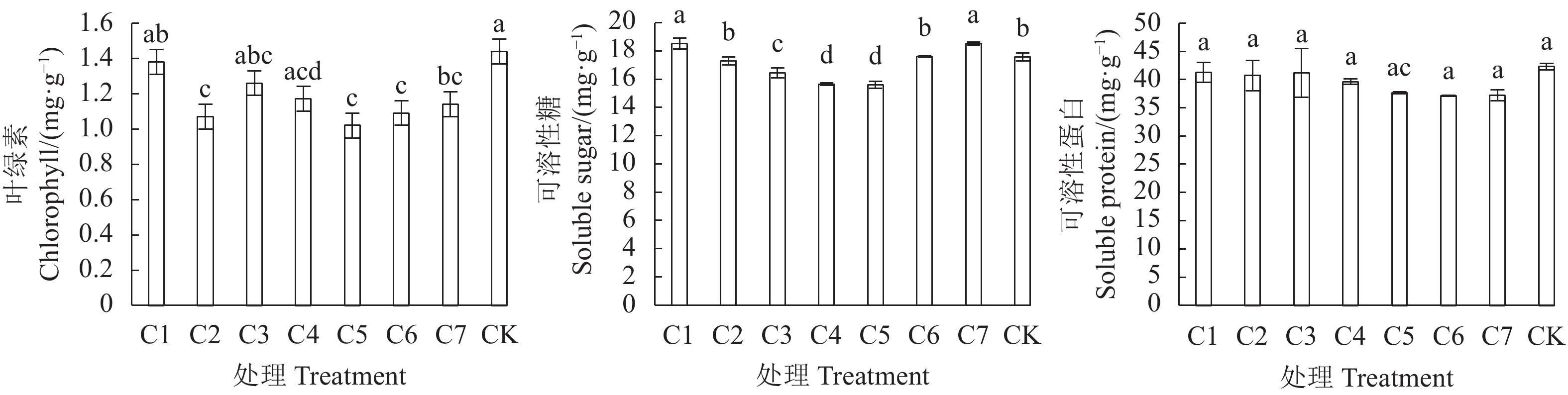
图 3 不同穴盘袋栽套袋时长金线莲生理指标
注:图中不同小写字母表示差异显著(P<0.05)。图4同。
Figure 3. Physiological indices on A. roburghii cultivated with varied bagging durations
Note: Data with different lowercase letters indicate significant difference at 0.05 level. Same for Fig. 4.
![]()
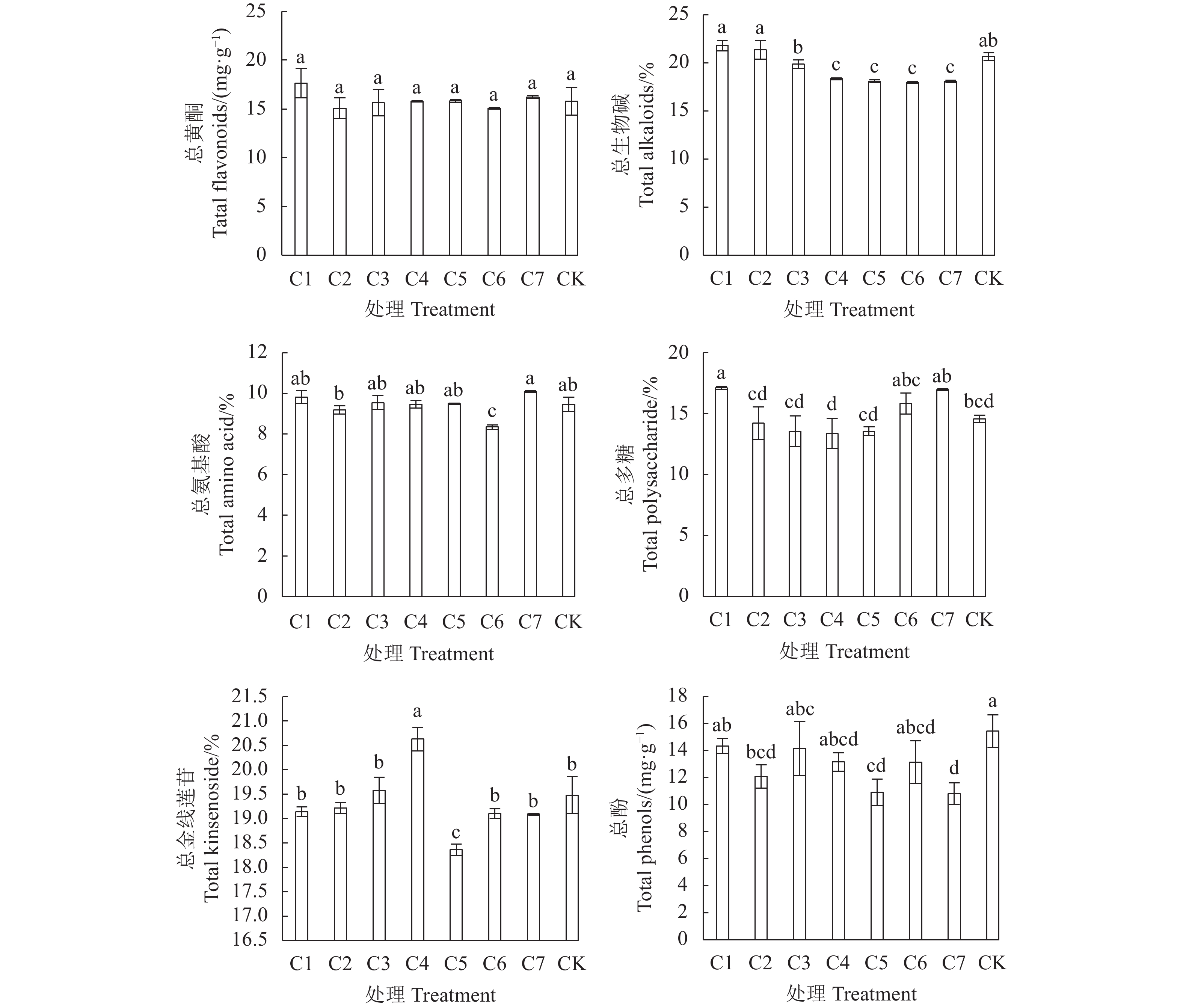
图 4 不同穴盘袋栽套袋时长金线莲活性成分图
Figure 4. Functional ingredients in A. roburghii cultivated with varied bagging durations
表 1 不同穴盘袋栽套袋时长金线莲平均单株鲜、干重及折干率
Table 1 Average fresh and dry weights and drying rates of A. roburghii cultivated with varied bagging durations
处理
Treatment平均单株鲜重
Average fresh
weight per plant/g平均单株干重
Average dry
weight per plant/g折干率
Discount
rate/%C1 5.14±0.41 b 0.99±0.07 ab 19.38±0.26 ab C2 4.75±0.11 c 0.89±0.03 abc 18.76±0.28 bc C3 4.07±0.13 d 0.75±0.02 abc 18.39±0.22 c C4 4.01±0.90 d 0.74±0.17 bc 18.58±0.38 c C5 3.94±1.07 d 0.68±0.19 c 17.27±0.22 d C6 5.78±0.47 a 1.00±0.08 ab 17.30±0.18 d C7 5.94±0.35 a 0.96±0.06 abc 16.26±0.15 e CK 5.29±0.37 b 1.04±0.06 a 19.73±0.20 a 注:同列数据后不同小写字母表示不同处理间差异显著(P<0.05)。
表2同。
Note: Data with different lowercase letters on same column indicate significant differences between treatments (P<0.05). Same for Table 2. 下载: 导出CSV
下载: 导出CSV
表 2 不同穴盘袋栽套袋时长金线莲生长指标
Table 2 Growth of A. roburghii cultivated with varied bagging durations
处理
Treatment株高
Plant
height/cm叶片数
Number of
leaves叶长
Leaf
length/cm叶宽
Leaf
width/cm茎节数
Number of
stems每节长
Length of
per section/cm根数
Root
number根长
Root
length/cmC1 9.12±0.17 d 5.00±0.17 c 3.31±0.73 a 2.25±0.40 a 4.00±0.13 a 2.46±0.06 d 4.60±0.89 a 4.26±2.84 a C2 9.30±0.03 cd 5.20±0.10 c 2.59±0.79 a 2.01±0.57 a 3.00±0.71 a 2.54±0.03 d 4.60±1.14 a 4.44±2.42 a C3 9.50±0.05 c 5.37±0.06 bc 2.94±0.69 a 2.06±0.34 a 4.00±0.23 a 2.56±0.05 cd 5.00±1.00 a 4.74±2.68 a C4 9.57±0.04 c 5.20±0.10 c 3.07±0.87 a 2.17±0.49 a 4.00±0.12 a 2.56±0.11 cd 4.60±0.89 a 5.44±2.51 a C5 10.14±0.13 b 5.73±0.25 ab 3.01±0.98 a 2.09±0.68 a 3.60±0.55 a 2.72±0.04 bc 4.40±0.89 a 5.28±1.71 a C6 10.51±0.03 a 6.00±0.26 a 3.72±0.9 a 2.72±0.63 a 3.80±0.45 a 2.83±0.09 b 4.80±0.45 a 5.58±1.95 a C7 10.62±0.04 a 6.10±0.20 a 4.14±0.75 a 2.93±0.42 a 4.00±0.71 a 3.14±0.02 a 5.20±0.84 a 8.34±0.74 a CK 8.31±0.2 e 4.40±0.10 d 2.85±0.99 a 2.10±0.52 a 3.40±0.55 a 2.39±0.02 d 4.40±1.34 a 6.32±0.94 a
下载: 导出CSV
-
[1] 洪琳, 邵清松, 周爱存, 等. 金线莲产业现状及可持续发展对策 [J]. 中国中药杂志, 2016, 41(3):553−558. HONG L, SHAO Q S, ZHOU A C, et al. Current status and sustainable development countermeasures of Anoectochilus roxburghii [J]. China Journal of Chinese Materia Medica, 2016, 41(3): 553−558.(in Chinese)
[2] 李秋静, 郑晓艳, 刘桂君, 等. 福建药用植物金线莲研究进展 [J]. 世界科学技术-中医药现代化, 2018, 20(8):1364−1372. LI Q J, ZHENG X Y, LIU G J, et al. Overview of the research on the medical herb of Fujian Province-Anoectochilus roxburghii [J]. Modernization of Traditional Chinese Medicine and Materia Medica-World Science and Technology, 2018, 20(8): 1364−1372.(in Chinese)
[3] 牛欢, 谢卓宓, 古力, 等. 林下种植密度和收获期对金线莲产量和品质的影响 [J]. 中国现代中药, 2018, 20(7):837−841,865. NIU H, XIE Z M, GU L, et al. Effects of planting density and harvesting stages for Anoectochilus roxburghii planted under forest on its yield and quality [J]. Modern Chinese Medicine, 2018, 20(7): 837−841,865.(in Chinese)
[4] 甘金佳, 毛玲莉, 黄容乐, 等. 不同栽培方式对金线莲生长状况和品质的影响 [J]. 中国农业科技导报, 2018, 20(7):130−136. GAN J J, MAO L L, HUANG R L, et al. Effects of different cultivation methods on the growth and quality of Anoectochilus roxburghii [J]. Journal of Agricultural Science and Technology, 2018, 20(7): 130−136.(in Chinese)
[5] 陈仁清. 密度和气候年景对林下仿野生栽培金线莲生长的影响 [J]. 安徽农学通报, 2019, 25(5):26−28,50. DOI: 10.3969/j.issn.1007-7731.2019.05.012 CHEN R Q. Effects of density and climate year on the growth of imitating wild cultivated Anoectochilus roxburghii under forest [J]. Anhui Agricultural Science Bulletin, 2019, 25(5): 26−28,50.(in Chinese) DOI: 10.3969/j.issn.1007-7731.2019.05.012
[6] 郭剑雄, 洪佰仲. 金线莲袋式优质高产有机栽培技术 [J]. 农技服务, 2017, 34(10):20−21. GUO J X, HONG B Z. The bag-type high-quality and high-yield organic cultivation technology of Anoectochilus roxburghii [J]. Agricultural Technology Service, 2017, 34(10): 20−21.(in Chinese)
[7] 吴纯清, 程昌凤, 唐小华. 水果套袋的研究现状 [J]. 西南园艺, 2002(S1):28−30,36. WU C Q, CHENG C F, TANG X H. Research status of fruit bagging [J]. Southwest Horticulture, 2002(S1): 28−30,36.(in Chinese)
[8] 朱俊斌. 一种带气孔结构的微型金线莲种植棚: CN203934426U[P]. 2014-11-12. [9] 郑连金, 马增强, 肖玉兰. 不同光照强度对台湾金线莲生长发育和次生代谢物合成的影响 [J]. 安徽农学通报, 2016, 22(16):25−26, 39. DOI: 10.3969/j.issn.1007-7731.2016.16.010 ZHENG L J, MA Z Q, XIAO Y L. The influence of different light intensities on the growth and development and the synthesis of secondary metabolites of aoectochilus formosanus [J]. Anhui Agricultural Science Bulletin, 2016, 22(16): 25−26, 39.(in Chinese) DOI: 10.3969/j.issn.1007-7731.2016.16.010
[10] 张超, 吴建国, 易骏, 等. HPLC-ELSD法测定三种植物基原金线莲的金线莲苷含量 [J]. 食品工业科技, 2017, 38(2):75−78. ZHANG C, WU J G, YI J, et al. Content determination of kinsenoside in Jin-Xian-Lian from three Anoectochilus species by HPLC-ELSD [J]. Science and Technology of Food Industry, 2017, 38(2): 75−78.(in Chinese)
[11] 魏翠华, 谢宇, 秦建彬, 等. 不同品种金线莲氨基酸和多糖含量的比较研究 [J]. 福建林业科技, 2016, 43(1):43−45,60. WEI C H, XIE Y, QIN J B, et al. Comparative study of amino acid and polysaccharide contents in different varieties of Anoectochilus roxburghii [J]. Journal of Fujian Forestry Science and Technology, 2016, 43(1): 43−45,60.(in Chinese)
[12] 孙加焱, 余红伟, 方锡平. 不同月龄林下仿野生栽培金线莲有效成分含量研究 [J]. 现代农业科技, 2020(11):66−67. DOI: 10.3969/j.issn.1007-5739.2020.11.036 SUN J Y, YU H W, FANG X P. Study on the Contents of Active Ingredients in Imitating Wild Cultivation of Anoectochilus roxburghii under Forests of Different Months [J]. Modern Agricultural Science and Technology, 2020(11): 66−67.(in Chinese) DOI: 10.3969/j.issn.1007-5739.2020.11.036
[13] ZUO L L, WANG Z Y, FAN Z L, et al. Evaluation of antioxidant and antiproliferative properties of three Actinidia (Actinidia kolomikta, Actinidia arguta, Actinidia chinensis) extracts in vitro [J]. International Journal of Molecular Sciences, 2012, 13(5): 5506−5518. DOI: 10.3390/ijms13055506
[14] 梅瑜, 邱道寿. 金线莲活性成分和分子鉴定的研究进展 [J]. 安徽农业科学, 2018, 46(9):29−33. DOI: 10.3969/j.issn.0517-6611.2018.09.009 MEI Y, QIU D S. Research progress on the active components and molecular identification of Anoectochilus sp. (wall. ) lindl [J]. Journal of Anhui Agricultural Sciences, 2018, 46(9): 29−33.(in Chinese) DOI: 10.3969/j.issn.0517-6611.2018.09.009
[15] 龚秀会, 许敏, 董鸿竹, 等. 不同基原金线莲植物的化学成分比较研究 [J]. 安徽农业科学, 2012, 40(36):17530−17531,17607. DOI: 10.3969/j.issn.0517-6611.2012.36.032 GONG X H, XU M, DONG H Z, et al. Comparative Study of the Chemical Constituents of Anoectochilus spp. from different origins [J]. Journal of Anhui Agricultural Sciences, 2012, 40(36): 17530−17531,17607.(in Chinese) DOI: 10.3969/j.issn.0517-6611.2012.36.032
[16] 赖曼娜, 林汝秀, 钟鸣. Box-Behnken设计优化金线莲总生物碱提取工艺研究 [J]. 中药材, 2017, 40(8):1910−1913. LAI M N, LIN R X, ZHONG M. Box-Behnken Design and Optimization of Extraction Process of Total Alkaloids from Anoectochilus roxburghii [J]. Journal of Chinese Medicinal Materials, 2017, 40(8): 1910−1913.(in Chinese)
[17] 邵玲, 梁广坚, 刘楠, 等. 广东肇庆地区金线莲低海拔林下栽培研究 [J]. 热带亚热带植物学报, 2017, 25(6):546−553. DOI: 10.11926/jtsb.3769 SHAO L, LIANG G J, LIU N, et al. Studies on cultivation under forest of Anoectochilus roxburghii at low altitude in Zhaoqing, Guangdong Province, China [J]. Journal of Tropical and Subtropical Botany, 2017, 25(6): 546−553.(in Chinese) DOI: 10.11926/jtsb.3769
[18] 朱冬梅, 朱添福, 邱光敏, 等. 福建省武平县金线莲种植气候条件 [J]. 农业工程, 2016, 6(5):132−133. DOI: 10.3969/j.issn.2095-1795.2016.05.040 ZHU D M, ZHU T F, QIU G M, et al. Climatic conditions of Anoectochilus roxburghii planting in Wuping County of Fujian Province [J]. Agricultural Engineering, 2016, 6(5): 132−133.(in Chinese) DOI: 10.3969/j.issn.2095-1795.2016.05.040
[19] 王大平, 杨玲. 套袋对黄花梨果实品质和贮藏性能的影响 [J]. 北方园艺, 2010(1):17−19. WANG D P, YANG L. Effect of bagging on quality and storage property of Huanghua pear [J]. Northern Horticulture, 2010(1): 17−19.(in Chinese)
[20] SHARMA R R, REDDY S V R, JHALEGAR M J. Pre-harvest fruit bagging: A useful approach for plant protection and improved post-harvest fruit quality-a review [J]. The Journal of Horticultural Science and Biotechnology, 2014, 89(2): 101−113. DOI: 10.1080/14620316.2014.11513055
[21] TEIXEIRA R, BOFF M I C, AMARANTE C V T D, et al. Effects of fruit bagging on pests and diseases control and on quality and maturity of ‘Fuji Suprema’ apples [J]. Bragantia, 2010, 70(3): 688−695.
[22] 吴世涛. 套袋对柚果实色泽变化及品质的影响[D]. 福州: 福建农林大学, 2019. WU S T. Effects of bagging on color change and quality of pummelo fruit[D]. Fuzhou: Fujian Agriculture and Forestry University, 2019. (in Chinese)
[23] 李章, 臧小平, 葛宇, 等. 套袋对芒果中甲基硫菌灵与吡虫啉农药残留的影响 [J]. 热带作物学报, 2017, 38(2):353−358. DOI: 10.3969/j.issn.1000-2561.2017.02.026 LI Z, ZANG X P, GE Y, et al. Effects of fruit bagging on residual of thiophanate-methyl and imidacloprid in mango fruit [J]. Chinese Journal of Tropical Crops, 2017, 38(2): 353−358.(in Chinese) DOI: 10.3969/j.issn.1000-2561.2017.02.026
[24] 武维华. 植物生理学[M]. 3版. 北京: 科学出版社, 2018. [25] 邱尔发, 陈存及, 董建文, 等. 毛竹种源新竹叶绿素含量动态变化及其相关关系研究 [J]. 江西农业大学学报(自然科学), 2002, 24(4):522−527. QIU E F, CHEN C J, DONG J W, et al. A study on chlorophyll content dynamics and relation ship of new bamboo of Phyllostachys heterocycla cv. pubescens from different provenances [J]. Acta Agriculturae Universitis Jiangxiensis, 2002, 24(4): 522−527.(in Chinese)
[26] 王艳. 设施条件下柑橘生物学特性及生理反应的初步研究[D]. 武汉: 华中农业大学, 2007. WANG Y. Preliminary study on biological characteristic and physiological reaction of Citrus in protected cultivate conditions[D]. Wuhan: Huazhong Agricultural University, 2007. (in Chinese)
[27] 牛欢, 韦坤华, 徐倩, 等. 不同光照度对金线莲生长、生理特性和药用成分的影响 [J]. 植物资源与环境学报, 2020, 29(1):26−36, 43. DOI: 10.3969/j.issn.1674-7895.2020.01.04 NIU H, WEI K H, XU Q, et al. Effects of different illuminances on growth, physiological characteristics, and medicinal components of Anoectochilus roxburghii [J]. Journal of Plant Resources and Environment, 2020, 29(1): 26−36, 43.(in Chinese) DOI: 10.3969/j.issn.1674-7895.2020.01.04
[28] 何碧珠, 邹双全, 刘江枫, 等. 光照强度与栽培模式对金线莲生长及品质影响 [J]. 中国现代中药, 2015, 17(12):1292−1295. HE B Z, ZOU S Q, LIU J F, et al. Effects of light intensity and cultivation mode on growth and quality of Anoectochilus roxburghii [J]. Modern Chinese Medicine, 2015, 17(12): 1292−1295.(in Chinese)
-
期刊类型引用(2)
1. 孙伟豪,刘亭,桑祎楠,杨政伟,张改生,宋瑜龙,张双喜. 宁夏引黄灌区春小麦品种(系)抗倒伏相关性状杂种优势及配合力分析. 中国农业科学. 2024(13): 2497-2508 .  百度学术
百度学术
2. 李忠金,付思远,陈萍萍,游月华,戴展峰,黄水明,彭玉林. 水稻籼粳中间型杂交组合若干性状的遗传效应分析. 福建农业学报. 2024(05): 522-531 . 本站查看
其他类型引用(1)

计量
- 文章访问数: 884
- HTML全文浏览量: 321
- PDF下载量: 32
- 被引次数: 3
