

Correlation Between Ectoenzyme Activity and Virulence against Planococcus citri of Metarhizium anisopliae F061
-
摘要: 室内测定台湾黑僵菌F061菌株的孢外酶活性及其对柑橘粉蚧的毒力,并通过线性回归分析探讨两者的相关性。结果表明:供试菌株的2种胞外酶活性均在1.00×108和1.00×107孢子·mL-12个处理时较高,其中蛋白酶活性达7.63 U·mL-1和7.61 U·mL-1,几丁质酶活性达5.45 U·mL-1和5.67 U·mL-1;供试菌株对柑橘粉蚧的LT50也在1.00×108和1.00×107孢子·mL-12个处理时较短,分别为4.98 d和5.39 d;线性回归分析表明,供试菌株对柑橘粉蚧的LT50与菌株蛋白酶活性呈显著负相关,与菌株几丁质酶活性呈中度负相关。可见,台湾黑僵菌F061菌株蛋白酶活性比几丁质酶活性强,在侵染柑橘粉蚧过程中起更直接的作用。Abstract: The ectoenzyme activity and the virulence against Planococcus citri of Metarhizium anisopliae F061 were determined in the laboratory. Their correlation was established using a linear regression analysis. The activities of two ectoenzymes from the bacteria were high.At concentrations of 1.00×108 and 1.00×107 conidia·mL-1, the protease activities of the spores were 7.63 U·mL-1 and 7.61 U·mL-1, respectively; while the chitinase activities, 5.45 U·mL-1 and 5.67 U·mL-1, respectively; and, the LT50 of the strain against P.citri, 4.98 d and 5.39 d, respectively. The LT50 inversely correlated significantly with the protease activity, but moderately with the chitinase activity. It appeared that the protease were stronger than the chitinase in M.anisopliae F061, and thus, played a more critical role in infecting P. citri.
-
Keywords:
- Metarhizium anisopliae /
- ectoenzyme /
- Planococcus citri /
- virulence /
- correlation
-
柑橘粉蚧Planococcus Citri又称桔臀纹粉蚧,属半翅目Hemiptera粉蚧科Pseudococcidae害虫,寄主广,在福建除危害柑橘类植物外,还可危害菠萝、柿、葡萄、石榴、草莓、龙眼、枇杷、橄榄等多种水果及多种观赏植物[1],引起落叶、落花和落果,分泌的蜜露还可诱发煤烟病,使为害加重,直接或间接影响了花果的品质和经济价值,不易防治,需要寻找一种较为安全、有效和经济的防控措施。目前,该虫的防控偏重于使用化学农药,易造成“3R”和药害问题[2-4]。生物防治是一种安全、有效、持久的控害方法,且虫生真菌挖掘利用已是害虫生防的重要发展方向之一[5]。台湾黑僵菌Metarhizium anisopliae是一种地理分布和寄主范围均比较广泛的虫生真菌,在台湾和国外已广泛用于防控介壳虫、金龟子、象甲、蝉类和蛾类等害虫[6];其F061菌株为台湾联发生物科技股份有限公司所研发,目前在福建省防控茶小绿叶蝉Empoasca pirisuga、茶丽纹象虫Myllocerinus aurolineatus、油茶毒蛾Euproctis pseudoconspersa等害虫取得良好的效果[7],但该菌株的孢外酶活性变化情况与其对柑橘粉蚧毒力的相关性未见报道,影响了该菌株在花果害虫防控领域的推广应用。本研究以台湾黑僵菌F061菌株为供试菌株,以柑橘粉蚧成虫为供试虫源,测定供试菌株的蛋白酶和几丁质酶等胞外酶活性及目标害虫感染该菌后的日死亡率,再通过线性回归分析探讨两者的相关性,以期为利用虫生真菌防控柑橘粉蚧等害虫提供技术支持。
1. 材料与方法
1.1 供试材料
供试台湾黑僵菌F061菌株母种由台湾联发生物科技股份有限公司提供,保存和活化均采用察氏培养基;供试柑橘粉蚧采集于福建省建瓯市东峰镇的桔柑园,在温度 (25±2)℃、相对湿度 (60±5)%、光周期L:D=12:12、光照强度 (10000±50) lx条件下饲养8~10代,取蜕皮3 d的成虫备用;供试试剂均为分析纯。
1.2 试验方法
1.2.1 菌株活化及菌液制备
将斜面保存的供试菌株接种到9 cm的察氏培养基平板上,置于温度 (25±1)℃、相对湿度 (90±5)%、光周期L:D=10:14、光照强度 (2000±50) lx的HGZ-150型光照培养箱内活化培养8 d;后加入5 mL含0.05%吐温-80的无菌水脱溶孢子,经3层脱脂纱布过滤得孢子原液;用纽鲍尔血细胞计数板在B203LEDTR型生物显微镜下统计孢子原液的孢子数并计算孢子浓度,再用无菌水将孢子原液稀释成1.00×108、1.00×107、1.00×106、1.00×105和1.00×104孢子·mL-1等5个浓度的供试菌液备用。
1.2.2 酶活诱导培养基制备
先将柑橘粉蚧成虫置于-80℃冰箱中速冻2 h,再置于60℃的DHG-9030型电热鼓风干燥箱中烘干6 h,后用玻璃研钵研磨成细粉末,虫尸粉量不低于2.0 g;再参考董晶[8]的方法称取K2HPO4 1 g、KCl 0.5 g、MgSO4·7H2O 0.5 g和FeSO4·7H2O 0.01 g溶于1 000 mL蒸馏水中,制成基本培养基;后称取柑橘粉蚧虫尸粉0.2 g作为唯一的碳氮源,溶于100 mL基本培养基中,置于121℃的LDZX-50FBS型不锈钢立式压力蒸汽灭菌器中灭菌20 min,冷却后制成含酶活诱导基质的液体培养基备用。
1.2.3 供试菌株孢外酶活性测定
蛋白酶活性测定:取1.2.1制备的5个浓度供试菌液各10 mL,参考董晶[8]的方法,分别接入到100 mL含1.2.2所制液体培养基的三角瓶中,后置于与1.2.1相同条件的ZWY-240型旋转式摇床上分别培养8 d,过滤得5份粗酶液备用;接着以酪蛋白为底物,加入0.05 mol·L-1、pH=8.5的Tris-HCl缓冲液配成1%(W/V) 的酪蛋白溶液,定容至100 mL备用;然后参考彭国良等[9]的方法,取10支试管,每份粗酶液分配2支试管,其中一支试管先加入1.0 mL的粗酶液和酪蛋白溶液摇匀,置于37℃水浴中反应30 min,再加入3 mL、0.4 mol·L-1的三氯乙酸溶液终止反应,另一支试管作为对照,则先加入1.0 mL待测粗酶液和3 mL、0.4 mol·L-1的三氯乙酸溶液摇匀,置于37℃水浴中反应30 min,再加入1.0 mL酪蛋白溶液摇匀,将2支试管内的反应液过滤,分别取1 mL滤液,均先加入5 mL、0.55 mol·L-1的碳酸钠溶液混匀,再加入1.0 mL福林酚试剂混匀,显色30 min,后用UV-1800PC型紫外可见分光光度计在680 nm处测吸光值,根据酪氨酸标准曲线计算8份粗酶液的蛋白酶活性,以每分钟催化分解酪蛋白生成1 μg酪氨酸的酶量为1个酶活单位。几丁质酶活性测定:取1.2.1制备的5个浓度供试菌液各10 mL,按董晶[8]的方法,分别接入100 mL含1.2.2所制液体培养基的三角瓶中,后置于与1.2.1相同条件的旋转式摇床上分别培养8 d,滤纸过滤得5份粗酶液;接着按彭国良等[9]的方法,取10支试管,每份粗酶液分配2支试管,其中一支试管加入0.5 mL待测粗酶液,另一支试管加入0.5 mL经100℃加热失活的粗酶液作为对照,再往2支试管中分别加入0.5 mL胶状几丁质,置37℃水浴中反应4 h,再用KDC-140HR型高速冷冻离心机8 500 r·min-1下离心5 min使反应停止,接着吸取0.2 mL上清液加入0.08 mL四硼酸钾溶液摇匀,在沸水浴中反应5 min,迅速用自来水冷却却室温,再加入1.2 mL、10%的二甲氨基苯甲醛试剂摇匀,在37℃下保温20 min,用自来水将反应液冷却至室温,后用紫外可见分光光度计在585 nm处测吸光值,根据N-乙酰氨基葡萄糖标准曲线计算8份粗酶液的几丁质酶活性,以每分钟催化分解几丁质生成1 μg N-乙酰氨基葡萄糖的酶量为1个酶活单位。
1.2.4 供试菌株对柑橘粉蚧的毒力测定
采用浸虫法测定。取1.2.1制备的5个浓度供试菌液,以0.05%吐温-80无菌水作空白对照,共6个处理,每个处理重复3次;再取直径9 cm的培养皿,依次铺入相同直径的花泥和滤纸各1片,花泥厚0.5 cm吸水6 mL,在滤纸上再放1片带叶柄的柑橘嫩叶,叶柄用湿棉花包裹并紧贴滤纸;接着用软毛笔刷往每个处理挑取供试成虫30头,浸泡10 s后挑到滤纸上,晾干2 min后再挑到叶片上,盖上培养皿;后将培养皿置于与1.2.1相同条件的光照培养箱内,每日14:00观察、记录死虫数,以死亡虫体长出菌丝计为有效感染,每次记录后挑除死亡虫体),8 d后统计试虫的死亡数并估测每株菌株的致死中时LT50。
1.3 数据统计与分析
在DPS 7.05数据处理软件上,利用DMRT检验法对供试菌株在不同孢子浓度的胞外酶活性进行多重比较;利用机率值分析法估测供试菌株对柑橘粉蚧成虫的LT50;利用线性回归分析法构建供试菌株胞外酶活性与其对柑橘粉蚧毒力的回归方程,分析两者的相关性。
2. 结果与分析
2.1 供试菌株的胞外酶活性
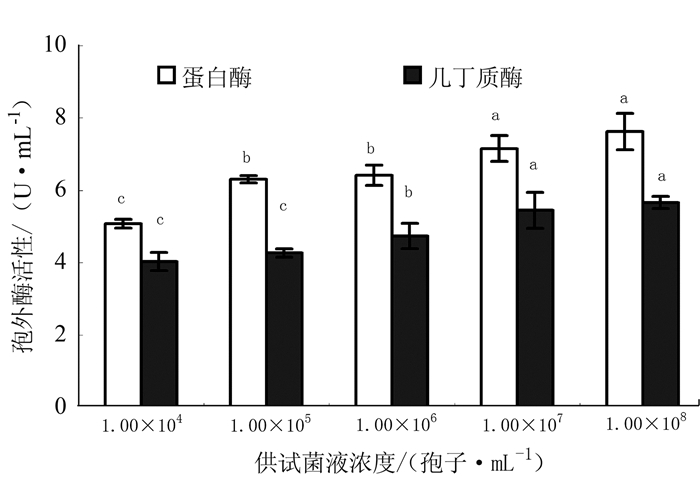
从图 1可知,供试菌株在不同孢子浓度时的2种胞外酶活性各不相同。蛋白酶活性,以1.00×108和1.00×107孢子·mL-12个处理较高,分别达7.63 U·mL-1和7.16 U·mL-1,显著高于其他3个处理;1.00×106和1.00×105孢子·mL-1处理位居其次,相互间差异不显著;1.00×104孢子·mL-1最低,仅为1.00×108和1.00×107孢子·mL-1 2个处理的66.58%和70.95%。几丁质酶活性,也以1.00×108和1.00×107孢子·mL-1两个处理较高,分别达5.67 U·mL-1和5.45 U·mL-1,显著高于其他3个处理;1.00×106孢子·mL-1处理位居其次;1.00×105和1.00×104孢子·mL-12个处理较低,与其他处理差异显著。
![]() 图 1 台湾黑僵菌F061菌株的胞外酶活性注:图中不同字母表示差异显著 (P<0.05)。Figure 1. Ectoenzyme activity of M. anisopliae F061
图 1 台湾黑僵菌F061菌株的胞外酶活性注:图中不同字母表示差异显著 (P<0.05)。Figure 1. Ectoenzyme activity of M. anisopliae F0612.2 供试菌株对柑橘粉蚧成虫的LT50
从表 1可知,供试菌株5个处理浓度对柑橘粉蚧成虫的LT50各不相同;1.00×108和1.00×107孢子·mL-12个处理较短,分别为4.98 d和5.39 d,相互间差异不显著,但显著短于其他3个处理;1.00×106和1.00×105孢子·mL-12个处理相对较长,相互间差异也不显著;1.00×104孢子·mL-1处理最长,分别达1.00×108和1.00×107孢子·mL-12个处理的1.93倍和1.79倍。
表 1 台湾黑僵菌F061菌株对柑橘粉蚧成虫的LT50Table 1. LT50 of M. anisopliae F061 against P. citri孢子浓度/
(孢子·mL-1)毒力回归方程 相关系数 LT50
/d95%置信区间
/d1.00×104 y=0.6256+4.4463x 0.9577 9.63 (7.31~12.70) a 1.00×105 y=0.5833+4.9733x 0.9710 7.73 (6.38~9.36) b 1.00×106 y=0.4428+5.4980x 0.9846 6.74 (5.95~7.64) b 1.00×107 y=0.5276+6.1150x 0.9826 5.39 (4.81~6.04) c 1.00×108 y=0.4795+6.4818x 0.9866 4.98 (4.53~5.48) c 注:表中同列不同字母表示差异显著 (P<0.05)。 2.3 供试菌株胞外酶活性与其对柑橘粉蚧毒力的相关性
通过线性回归分析,得到供试菌株蛋白酶活性 (x1) 和几丁质酶活性 (x2) 与其对柑橘粉蚧成虫LT50(y) 的回归方程y=19.5996-1.3168x1-0.8527x2,方程显著性检验达显著水平 (P=0.02<0.05);同时由方程变量的偏相关分析可知,供试菌株对柑橘粉蚧成虫的LT50与菌株蛋白酶活性呈显著负相关 (r=-0.8713,P=0.03<0.05),与供试菌株几丁质酶活性变化呈中度负相关 (-0.5000≥r=-0.6469>-0.8000,P=0.55>0.05)。
3. 讨论与结论
虫生真菌在侵染昆虫的过程中,蛋白酶、几丁质酶等胞外酶在降解昆虫体壁方面发挥极其重要的作用[8-11],但有关胞外酶活性与毒力相关性的认识目前尚未达成一致[12]。李会平等[11]和雷妍圆等[12]分别发现球孢白僵菌Beauveria bassiana对桑天牛Apriona germari和小菜蛾Plutella xylostella的毒力与菌株蛋白酶和几丁质酶活性均呈显著相关;而冯明光[13]认为球孢白僵菌对血黑蝗Melanoplus sanguinipes的毒力与菌株蛋白酶活性呈显著相关,但与几丁质酶活性是否具相同相关性尚存争议。本研究测定了台湾黑僵菌F061菌株在不同孢子浓度时的蛋白酶和几丁质酶活性,发现2种胞外酶活性均在1.00×108和1.00×107孢子·mL-12个处理时较高,但蛋白酶活性强于几丁质酶活性;同时也比较了台湾黑僵菌F061菌株在不同孢子浓度时对柑橘粉蚧成虫的毒力,发现供试菌株LT50也在1.00×108和1.00×107孢子·mL-12个处理时较高;通过对供试菌株胞外酶活性与其对柑橘粉蚧毒力的线性回归分析发现,供试菌株对柑橘粉蚧的毒力与菌株蛋白酶活性呈显著相关,与菌株几丁质酶活性呈中度相关,这与前人的部分研究成果一致[11-13]。可见,台湾黑僵菌F061菌株蛋白酶活性比几丁质酶活性强,在侵染柑橘粉蚧过程中起更直接的作用。
虫生真菌对寄主的致病和致死是一个复杂的生理生化过程,是菌株侵染与寄主防御等多方因子共同作用的结果。本研究虽初步明确了台湾黑僵菌F061菌株胞外酶活性与其对柑橘粉蚧毒力的相关性,但对柑橘粉蚧受侵染后的虫体组织器官病理变化情况及血淋巴中血细胞数量、蛋白质和氨基酸浓度、保护酶和解毒酶活性的变化等防御反应情况尚不清楚,因此今后有必要开展此方面的相关研究,同时进一步明确供试菌株的田间防效,以期为利用虫生真菌防控介壳虫等害虫提供技术支撑。
-
![]()
图 1 台湾黑僵菌F061菌株的胞外酶活性
注:图中不同字母表示差异显著 (P<0.05)。
Figure 1. Ectoenzyme activity of M. anisopliae F061
表 1 台湾黑僵菌F061菌株对柑橘粉蚧成虫的LT50
Table 1 LT50 of M. anisopliae F061 against P. citri
孢子浓度/
(孢子·mL-1)毒力回归方程 相关系数 LT50
/d95%置信区间
/d1.00×104 y=0.6256+4.4463x 0.9577 9.63 (7.31~12.70) a 1.00×105 y=0.5833+4.9733x 0.9710 7.73 (6.38~9.36) b 1.00×106 y=0.4428+5.4980x 0.9846 6.74 (5.95~7.64) b 1.00×107 y=0.5276+6.1150x 0.9826 5.39 (4.81~6.04) c 1.00×108 y=0.4795+6.4818x 0.9866 4.98 (4.53~5.48) c 注:表中同列不同字母表示差异显著 (P<0.05)。  下载: 导出CSV
下载: 导出CSV
-
[1] 黄邦侃.福建昆虫志第2卷[M].福州:福建科学技术出版社, 1999:675-676. [2] 王进强, 李国华. 蚧虫防治研究进展[C]//中国植物保护学会2009年学术年会论文集, 中国植物保护学会, 2009: 1051. [3] 郭莹, 刘长明, 吴梅香, 等.防治橘臀纹粉蚧药剂的室内筛选[J].武夷科学, 2012, 28(1):102-105. http://www.cnki.com.cn/Article/CJFDTOTAL-WYKX201200018.htm [4] 郭俊, 赖新朴, 高俊燕, 等.云南柠檬园橘臀纹粉蚧的发生及防治初探[J].植物保护, 2014, 40(4):157-160. http://www.cnki.com.cn/Article/CJFDTOTAL-ZWBH201404032.htm [5] 王联德, 尤民生, 黄建, 等.虫生真菌多样性及其在害虫生物防治中的作用[J].江西农业大学学报, 2010, 32(5):920-927. http://www.cnki.com.cn/Article/CJFDTOTAL-JXND201005015.htm [6] 林木一. Metarhizium anisopliae (黑殭菌)——生物制剂应用与实习[D]. 台湾省云林县: 虎尾科技大学, 2011. [7] 林金璇, 蓝炎阳, 陈毅勇, 等.台湾黑僵菌在茶叶生物防治上的应用[J].北京农业, 2014, (12):138-139. http://www.cnki.com.cn/Article/CJFDTOTAL-BJNY201412105.htm [8] 董晶, 谢映平, 刘卫敏, 等.病原真菌降解两种蚧虫体壁过程中胞外酶作用[J].中国农业科学, 2015, 48(5):889-899. http://www.cnki.com.cn/Article/CJFDTOTAL-ZNYK201505007.htm [9] 彭国良. 蜡蚧轮枝菌入侵蚧虫过程中胞外酶作用及提高酶活性的研究[D]. 太原: 山西大学, 2009. [10] 韩珍珍. 蜡蚧霉对蚧虫的致病力及组织病理学研究[D]. 太原: 山西大学, 2010. [11] 李会平, 黄大庄, 王志阏, 等.不同基质传代白僵菌菌株胞外蛋白酶和几丁质酶产生水平与菌株对桑天牛幼虫毒力的关系[J].蚕业科学, 2008, 34(3):521-524. http://www.cnki.com.cn/Article/CJFDTOTAL-CYKE200803025.htm [12] 雷妍圆, 吕利华, 何余容, 等.球孢白僵菌生物学特性与其对小菜蛾致病力相关性分析[J].中国生物防治, 2010, 26(2):143-148. http://www.cnki.com.cn/Article/CJFDTOTAL-ZSWF201002006.htm [13] 冯明光.胞外蛋白酶和脂酶活性作为球孢白僵菌毒力指标的可靠性分析[J].微生物学报, 1998, 38(6):461-467. http://www.cnki.com.cn/Article/CJFDTOTAL-WSXB199806008.htm -
期刊类型引用(2)
1. 黄鹏, 陈汉鑫, 姚锦爱, 黄建成, 余德亿. 金龟子绿僵菌对石蒜绵粉蚧的室内毒力与防治效果. 中国生物防治学报. 2019(06): 884-890 .  百度学术
百度学术
2. 黄鹏, 姚锦爱, 余德亿. 金龟子绿僵菌FM-03生物学特性及其对柑橘粉蚧的侵染. 中国生物防治学报. 2018(06): 858-865 . 百度学术
其他类型引用(0)
计量
- 文章访问数: 1453
- HTML全文浏览量: 185
- PDF下载量: 247
- 被引次数: 2
