

Effects of Arbuscular Mycorrhizal Fungi and Organic Fertilizer on Key Microbial Carbon-cycle Genes in Rhizosphere Soil at Sweet Corn Field
-
摘要:目的 研究丛枝菌根真菌(Arbuscular mycorrhizal fungi, AMF)与有机肥配施对甜玉米根际土壤微生物群落碳循环功能基因的影响,揭示AMF与有机肥配施对甜玉米根际土壤碳循环的生物学机制,为AMF配施有机肥在农田土壤碳转化方面的应用提供理论参考。方法 设置7个处理:(1)CK,不施氮肥;(2)OF,优化施肥;(3)ORF10,有机肥替代10%化学氮肥;(4)ORF20,有机肥替代20%化学氮肥;(5)ORF10+AMF,有机肥替代10%化学氮肥+变形球囊霉菌(Glomus versiforme);(6)ORF20+AMF,有机肥替代20%化学氮肥+变形球囊霉菌(Glomus versiforme);(7)CK+AMF,不施氮肥+变形球囊霉菌(Glomus versiforme);每个处理3个重复。利用基因芯片(GeoChip5.0)技术,对7个处理甜玉米根际土壤关键碳循环功能基因进行分析。结果 AMF与有机肥配施对甜玉米产量具有显著的增产效果,在CK、ORF10、ORF20施肥水平下增施变形球囊霉菌后,甜玉米的鲜苞产量分别较不增施变形球囊霉的处理提升32.6%、8.6%、8.9%。测序结果显示,AMF与有机肥配施显著改变了甜玉米根际土壤微生物碳循环功能基因的结构特征。在关键碳循环基因水平上,与碳循环有关的碳分解、碳固定和甲烷代谢3个过程总体上表现为CK+AMF与ORF20+AMF两个处理的基因相对信号强度均高于其他处理。关键碳循环功能基因与环境因子的冗余分析显示,土壤呼吸、全氮、pH、全钾、有机质、速效磷、全磷等土壤养分是影响碳循环功能基因的主要因素。结论 AMF与有机肥配施能显著提高甜玉米的鲜苞产量,同时也改变了甜玉米根际土壤碳循环功能基因的结构特征,不同程度提高了碳分解、碳固定、甲烷代谢等过程的功能基因的相对信号强度。Abstract:Objective Effect of arbuscular mycorrhizal fungi (AMF) and organic fertilizer applied in sweet corn field on the microbial genes relating to the carbon (C) cycling in the rhizosphere was studied to decipher the biological mechanism and the soil C-transformation.Method Seven treatments with triplicates each were applied on the sweet corn fields including (1) no N fertilizer (CK), (2) optimized fertilization (OF), (3) organic N fertilizer to replace 10% of chemical N fertilizer (ORF10), (4) organic N fertilizer to replace 20% of chemical N fertilizer (ORF20), (5) ORF10 with added Glomus versiforme (ORF10+AMF), (6) ORF20 with added G. versiforme (ORF20+AMF), and (7) CK with added G. versiforme (CK+AMF). Genes related to C-cycling in the treated rhizosphere soils were analyzed using GeoChip 5.0 technology.Result Addition of AMF in fertilizing the sweet corn plants significantly increased the yield. By adding AMF to CK, ORF10, and ORF20, the treatments increased the number of fresh buds on the plants by 32.6%, 8.6%, and 8.9%, respectively. The results of gene sequencing on the soil samples showed that the AMF/organic fertilizer combinations significantly altered the structure of the microbial C-cycle genes. The signal strength of key functional genes associated with C-cycling, such as C-decomposition, C-fixation, and methane metabolism, were generally stronger under CK+AMF and ORF20+AMF than the other treatments. According to the redundancy analysis, the respiration, total nitrogen, pH, total potassium, organic matter, available phosphorus, and total phosphorus in rhizosphere soil were the major environmental factors affecting the functions of the C-cycle-related genes.Conclusion The application of organic fertilizer and G. versiforme in sweet corn field significantly increased the fresh bud count on the plants. It changed the structure of the microbial C-cycle genes in rhizosphere soil positively affecting the decomposition and fixation of C as well as the methane metabolism of the ecosystem.
-
Keywords:
- Sweet corn /
- arbuscular mycorrhizal fungi /
- rhizosphere soil /
- soil microbe /
- functional genes /
- carbon-cycle
-
0. 引言
【研究意义】避雨栽培是目前常用的一种梨树种植方式,具有减少花期雨季落花、提高坐果率、缩短果实成熟时间、提高果实品质和减少病虫害等优势[1-3],而避雨栽培改变果园微域环境,特别是土壤环境,因而研究其对根际土壤细菌群落的影响,对完善避雨栽培下土壤管理措施具有重要意义。【前人研究进展】近些年来随着梨树种植的增加,梨树病虫害和恶劣的自然环境对梨品质和产量的相关报道越来越多[4-6]。为减少病虫害以及自然灾害对梨品质和产量的影响,避雨栽培模式在梨树种植的应用越来越多。避雨栽培下梨锈病和梨黑斑病的发病指数要明显小于露天栽培[7],且改善梨的外观,果实可溶性固形物和可溶性总糖含量都分别提高,可滴定酸含量和果实硬度都显著下降[8]。避雨栽培对梨品质和产量的研究相对较多,但避雨处理对梨树土壤的影响等研究相对较少,陈小明等[9]研究发现避雨栽培会减小梨树根际土壤酶活性。植物根际有一个复杂的微生物群落,它直接影响着植物的生长、健康和发育,根际微生物的群落结构受植物根际分泌物和土壤环境的影响[10-11]。【本研究切入点】避雨措施会影响梨树的生长,这势必会改变梨树的根际土壤环境,但对于避雨栽培下梨树根际土壤微生物的影响还未见报道。【拟解决的关键问题】本研究以同一梨园中避雨栽培和露地栽培的梨树为研究对象,测定梨果实品质、土壤理化性质和土壤微生物多样性,比较不同模式下梨树根际土壤根际微生物的变化与果实品质和土壤化学性质变化的相关性。为制定不同栽培模式下定向调控土壤微环境、提高水果品质和保护土壤环境等措施提供研究基础。
1. 材料和方法
1.1 试验地点
试验果园为福建省建宁县溪口镇枧头村67 hm2(约40000棵)30年生翠冠梨山地果园(东经116°48′34″,北纬26°50′38″),其中11.3 hm2(约6800棵)从2013年开始采用盖棚避雨栽培(TYGH),其余为无盖棚露地栽培(TY)。
1.2 梨果实品质分析
2018年7月14日进行梨的果实取样,取样方法为,按照5点取样法,在避雨栽培区和露地栽培区各随机选取5棵梨树。在每棵梨树的上层和下层的不同方位按东、西、南、北4个方位上,代表性的采4个果实,每棵树共采8个果实,每个处理共采40个。样品放入冰盒保存,当天带回实验室进行果实品质分析。
将带回实验室的果实,每个处理随机选择10个果,去皮后取中部果肉切碎混匀,重复4次。果实品质分析的测定参照中华人民共和国食品安全国家标准测试。测试方法如下:蔗糖含量测定采用《食品中果糖、葡萄糖、蔗糖、麦芽糖、乳糖的测定》(GB/T 5009.8—2016)[12],还原糖含量测定采用《食品中还原糖的测定》(GB/T 5009.7—2016)[13],抗坏血酸含量测定采用《食品中抗坏血酸的测定》(GB/T 5009.86—2016)[14],总酸含量测定采用《食品中总酸的测定》(GBT12456—2008)[15]。
1.3 土壤取样
随机选取避雨栽培的5株健壮梨树,同时在露地栽培区也随机选取5株健壮梨树,在距离树干1 m处,用铁铲挖10~20 cm深达梨树根,将树根取出,将根表面土壤刷下,然后把所有根系上刷下来的土壤剔除树根,充分混匀,每棵取500 g左右,共计2500 g。带回实验室保存于冰箱(−4 ℃)备用。
1.4 土壤化学测定
土壤样本取部分进行自然风干,碾碎,过2 mm筛,按四分法取样,参照《土壤农业化学分析方法》方法[16],测定土壤的pH值和全氮、全磷、全钾、速效氮、速效磷、速效钾以及有机质含量和含水量,所有指标测定均重复3次。具体如下:
1.4.1 土壤全氮和速效氮测定
速效氮含量包括测定的铵态氮和硝态氮含量的总和,铵态氮含量采用奈氏试剂法进行测定;硝态氮含量采用紫外分光光度法进行测定。全氮测定时先采用重铬酸钾硫酸消解法进行消解,再采用凯氏定氮法进行测定。
1.4.2 土壤全磷、全钾、速效磷和速效钾测定
用NaHCO3浸提土壤样品后采用钼锑抗比色法进行速效磷含量测定。利用NH4OAc浸提土壤,然后采用火焰光度法进行速效钾含量测定。全磷和全钾测定时先采用H2SO4-HClO4法进行消煮分解,再采用测定速效磷和速效钾的方法进行测定。
1.4.3 土壤有机质、pH和含水量测定
采用重铬酸钾-硫酸亚铁滴定法进行土壤有机质含量测定。
pH值测定简述为:取10 g土于250 mL烧杯中,加入25 mL无二氧化碳水,充分搅拌后平衡30 min。取上层清水,将校正好的pH计电极放入测定pH。
含水量测定简述为:先称取铝盒重量a,记录后加入新鲜土壤,记录铝盒和土壤的总重量m,放入烘箱100 ℃,24 h,后取出放在干燥器中冷却,称量铝盒和土壤的总重量b。土壤含水率=[(b−a)/(m−a)]×100%。
1.5 土壤DNA提取和测序
土壤总DNA的提取采用MoBio PowerSoil DNA isolation kit(Carlsbad, CA, USA)。利用1%琼脂糖凝胶电泳检测和分光光度法对提取到的DNA样品进行质量检测。将检测合格的样品于−20 ℃保存备用。采用338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)引物对细菌16S rRNA基因的V3-V4高变区进行扩增[17]。PCR反应体系为25μL,包括12.5 μL 2× Taq PCR MasterMix,3 μL BSA(2 ng·μL-1), 2 Primer(5 μM), 2 μL模板DNA,和5.5 μL ddH2O。反应参数:95 ℃预变性5 min;95 ℃变性45 s,55 ℃退火50 s,72 ℃延伸45 s,32个循环;72 ℃延伸10 min[18]。每个样品重复3次,将同一样本的PCR产物混合后用2%琼脂糖凝胶电泳检测,使用QIAquick Gel Extraction Kit(QIAGEN, Germany)凝胶回收试剂盒切胶回收PCR产物,Tris_HCl洗脱;利用RT-PCR进行定量,PCR产物送北京奥维森基因公司进行测序。
1.6 生物信息学分析
通过Illumina MiSeq平台进行Paired-end测序,下机数据经过QIIME(v1.8.0)软件过滤、拼接、去除嵌合体,去除打分低(≤20),碱基模糊,引物错配或测序长度小于150 bp的序列[19, 20]。根据barcodes规类各处理组序列信息聚类为用于物种分类的OUT(Operational Taxonomic Units),OTU相似性设置为97%(Edgar,2013)[21]。对比silva数据库,得到每个OTU对应的物种分类信息[22]。再利用Mothur软件(version 1.31.2)进行α多样分析(包括Shannon和Chao1等2个指数)[23]。基于Weighted Unifrace距离,使用R(v3.1.1)软件包的pheatmap进行聚类分析[24]。经过UniFrac算法利用系统进化的信息来比较样品间物种群落差异,并进行Beta多样性(Beta diversity)分析[25]。利用LEfSe分析进行不同处理根际细菌在丰度上有显著差异的细菌[26]。
1.7 数据分析
使用DPS软件分析数据,采用LSD法(最小显著差法)进行显著性差异分析,IBM SPSS Statistics 19.0进行数据的相关性分析,Umetrics SIMCA 13.0软件进行数据的主成分分析,并采用OriginLab OriginPro 2018进行绘图。
2. 结果与分析
2.1 梨树果实品质分析
梨果实品质测定结果表明,梨树避雨处理的果实蔗糖、还原糖含量要显著高于露地处理的,分别提高1.83倍和1.08倍,同时维生素C和总酸含量要显著低于露地处理的,分别减少了1.23倍和1.20倍(表1)。避雨处理果实的糖酸比要比露地处理的高19.45。可见避雨处理有利于提高果实糖含量和降低酸含量,提高果实的糖酸比,从而有可能改善风味品质。
表 1 不同栽培模式下梨果实品质Table 1. The pear fruit qualities under different cultivation modes样品
Samples蔗糖
Sucrose/g·hg−1还原糖
Reducing sugar/g·hg−1维生素C
Vitamin C/g·hg−1总酸
Total acid/g·kg−1糖酸比
Ratio of sugar to acidTYGH 1.28±0.01 a 7.17±0.01 a 6.18±0.05 b 1.20±0.01 b 70.42 TY 0.70±0.03 b 6.64±0.12 b 6.97±0.07 a 1.44±0.01 a 50.97 注:TYGH--避雨栽培, TY--露地栽培(图1~3同);小写字母表示P<0.05水平上的显著性(表2~3同)。
Note: “TYGH” represent rain-shelter cultivation, “TY” represent open field cultivation(The same as Fig.1-3). The lowercase represent significant difference at P<0.05 levels(The same as table 2-3.2.2 梨园土壤化学性质
土壤化学性质测定结果表明,在避雨处理的土壤速效磷、速效钾、有机质含量和含水量都要显著低于露地处理,全钾、速效氮含量和pH值都与露地处理没有显著差异,全氮含量显著高于露地处理(表2)。可见露地处理利于土壤速效磷、钾含量、有机质和含水量的提高,可能对果实的产量有重要作用。
表 2 不同栽培模式下梨树根际土壤化学性质Table 2. Chemical properties of soils from different treatment plots指标 Index TYGH TY 全钾 Total potassium/(g·kg−1) 5.75±0.14 a 5.88±0.20 a 全氮 Total nitrogen/(g·kg−1) 7.73±0.37 a 4.20±0.15 b 全磷 Total phosphorus/(g·kg−1) 0.066±0.004 a 0.090±0.015 a 速效氮 Available nitrogen/(mg·kg−1) 21.47±2.13 a 18.96±1.47 a 速效磷 Available phosphorus/(mg·kg−1) 87.74±7.11 b 165.38±17.66 a 速效钾 Available potassium/(mg·kg−1) 165.65±2.60 b 229.74±5.54 a 有机质 Organic matter/% 7.64±1.68 b 16.16±0.38 a pH 5.05±0.03 a 4.92±0.11 a 含水量 Water content/% 14.33±0.88 b 20.33±1.76 a 2.3 梨园土壤微生物测序分析
2.3.1 微生物序列分析
对梨树根际土壤微生物测序结果表明,避雨处理的RT(Raw_tags)数量和CT(Clean_tags)要显著低于露地处理,但最终的OUT数目要显著高于露地处理(表3)。说明避雨处理的梨树根际土壤微生物数量更多。
表 3 梨树根际土壤微生物序列拼接结果和OUT数目统计Table 3. Sequence alignment and OUT number statistics of pear rhizosphere soil microorganism样品 Samples RT CT OTUs TYGH 40774±3196 b 40441±3164 b 2009±28 a TY 44728±2854 a 44328±2845 a 1830±20 b 注:RT(Raw_tags):经过过滤低质量的fastq数据拼接得到的结果,CT(Clean_tags):进一步对拼接结果去除嵌合体、短序列后得到的结果, OTUs为样本最终得到的平均OTU数目。
Note: RT is the result of merging the filtered low-quality fastq data, CT is the result of further removing chimera and short sequences from the merging result, and OTUs is the average number of OTUs of the samples.2.3.2 梨园土壤微生物α多样性和丰富度
避雨处理的梨树根际土壤在门水平的细菌群落主要以变形菌门(Proteobacteria
)、酸杆菌门(Acidobacteria)、绿弯菌门(Chloroflexi)和放线菌门(Actinobacteria)为主(图1-A);在属水平的细菌群落主要以不可培养细菌(uncultured bacterium)、不可培养的森林土壤细菌(uncultured forest soil bacterium)和不可培养的变形菌(uncultured proteobacterium)为主(图1-B)。避雨处理的梨树根际土壤细菌群落的平均Shannon指数(香农指数)和Chao1都要大于露地处理的,避雨处理显著提高土壤细菌的多样性(P<0.05)。可见,避雨处理可以提高梨树根际土壤细菌群落的丰富度和多样性(图2)。 ![]() 图 1 不同栽培模式下梨树根际土壤细菌群落的门级(A)和属级(B)分类组成Figure 1. Taxonomic composition of bacterial communities in the rhizosphere soil of pear trees at the phylum level and genus level under different cultivation modes
图 1 不同栽培模式下梨树根际土壤细菌群落的门级(A)和属级(B)分类组成Figure 1. Taxonomic composition of bacterial communities in the rhizosphere soil of pear trees at the phylum level and genus level under different cultivation modes![]() 图 2 不同栽培模式下梨树根际土壤细菌群落的α多样性(香农指数A)和丰富度(B)Figure 2. The alpha diversity(Shannon index, A)and abundance(Chao1, B)of rhizosphere soil bacteria of pear trees under different cultivation modes.
图 2 不同栽培模式下梨树根际土壤细菌群落的α多样性(香农指数A)和丰富度(B)Figure 2. The alpha diversity(Shannon index, A)and abundance(Chao1, B)of rhizosphere soil bacteria of pear trees under different cultivation modes.2.3.3 梨园土壤微生物β多样性
PCA分析结果表明,主成分1可以很好地区分避雨处理和露地处理的梨树根际土壤细菌群落,主成分1解释变量方差的74.31%(图3)。
![]() 图 3 不同栽培模式下梨树根际土壤细菌群落主成分分析Figure 3. rincipal component analysis of bacterial community in pear rhizosphere soil under different cultivation patterns
图 3 不同栽培模式下梨树根际土壤细菌群落主成分分析Figure 3. rincipal component analysis of bacterial community in pear rhizosphere soil under different cultivation patterns2.3.4 梨园土壤微生物分类构成
从图4可以看出,避雨处理会使梨根际微生物产生差异。避雨处理的梨树根际微生物Gemmatimonadales、Chloroflexi、Nitrosomonadales、Thermoleophilia、Sphingomonadaceae、Streptomyces、Candidatus Koribacter、Hyphomicrobiaceae、Micrococcales、Micromonosporales、Acidimicrobiales、Corynebacterium、Anaerolinea的数量要显著大于露地处理。露地处理梨树根际微生物Solibacterales、Actinospicaceae、Corynebacteriaceae、Acidothemceae、Frankiales、Chitinophagaceae、Sphingobacteriales、Thermogemmatisporales、Roseiarcaceae、Acetobacteraceae、Rhodospirillales、Alcaligenaceae、Burkholderiaceae、Polyangiaceae、Myxococcales、Deltaproteobacteria、Xanthomonadaceae的数量要显著大于避雨处理。
![]() 图 4 不同栽培模式下梨树根际土壤细菌群落丰度变化Figure 4. Bacterial taxa with different abundance changes in rhizosphere soil of pear trees under different cultivation modes
图 4 不同栽培模式下梨树根际土壤细菌群落丰度变化Figure 4. Bacterial taxa with different abundance changes in rhizosphere soil of pear trees under different cultivation modes2.4 相关性分析
梨树不同管理处理的根际土壤微生物在属水平与土壤理化性质相关性结果表明(图5-A),主成分1解释了变量方差的87.41%,主成分2解释了变量方差的10.26%,梨树根际属水平微生物差异相关系数绝对值最高的是土壤速效钾含量。与露地处理根际属水平微生物差异相关系数最高的是土壤速效钾含量,而避雨处理梨树根际属水平微生物差异相关系数绝对值最高的则是土壤总氮含量。说明避雨处理对微生物属水平影响最大的是土壤氮素水平。果实的蔗糖含量与避雨处理梨树根际属水平微生物差异相关系数最高,果实的总酸含量与露地处理梨树根际属水平微生物差异相关系数最高(图5-B)。说明避雨处理可能通过影响土壤微生物来影响果实品质。
![]() 图 5 不同栽培模式下梨树根际土壤细菌群落的属级分类下与土壤理化性质(A)和梨品质(B)的相关性分析注:TolN—土壤总氮,TolP—土壤总磷,TolK—土壤总钾,AvaN—速效氮,AvaP—速效磷,AvaK—速效钾,OC—有机质含量,WC—含水量,pH—pH值,Sug—蔗糖含量,还原糖含量,TolA—总酸,Vc—维生素C。Figure 5. Correlation analysis between bacterial communities in the rhizosphere soil of pear trees at the genus level and the soil physio-chemical indexes(A)and fruit qualities under different cultivation modesNote: TolN—Soil total nitrogen, TolP—Soil total phosphorus, TolK—Soil total potassium, AvaN—Rapid available nitrogen, AvaP—Rapid available phosphorus, AvaK—Rapid available potassium, OC—Organic content, WC—Water content, pH—Acidity or alkalinity gradients, Sug—Sucrose content, RS—Reducing sugar content, TolA—Total acid, Vc— Vitamin C.
图 5 不同栽培模式下梨树根际土壤细菌群落的属级分类下与土壤理化性质(A)和梨品质(B)的相关性分析注:TolN—土壤总氮,TolP—土壤总磷,TolK—土壤总钾,AvaN—速效氮,AvaP—速效磷,AvaK—速效钾,OC—有机质含量,WC—含水量,pH—pH值,Sug—蔗糖含量,还原糖含量,TolA—总酸,Vc—维生素C。Figure 5. Correlation analysis between bacterial communities in the rhizosphere soil of pear trees at the genus level and the soil physio-chemical indexes(A)and fruit qualities under different cultivation modesNote: TolN—Soil total nitrogen, TolP—Soil total phosphorus, TolK—Soil total potassium, AvaN—Rapid available nitrogen, AvaP—Rapid available phosphorus, AvaK—Rapid available potassium, OC—Organic content, WC—Water content, pH—Acidity or alkalinity gradients, Sug—Sucrose content, RS—Reducing sugar content, TolA—Total acid, Vc— Vitamin C.3. 讨论
避雨设施栽培的应用是减少作物病虫害、提高作物产量和品质的一个重要措施[3, 27-29]。本研究发现山地避雨处理会提高梨的甜度,同时减少梨的总酸含量,该结果与前人的研究结果一致[3, 8, 30-32];陈小明等[9]对田地避雨栽培研究表明,田地避雨栽培并没有提高梨果实总的含糖量,但酸性物质含量有所增加,这与本研究结果不同,可能是由于田地与山地立地条件不同,土壤水肥养分积累及消耗能力不同。相对于田地,由于山地早晚温差大,紫外线强,梨树生长过程中应激响应大,梨树吸收矿质营养元素的效率更低,导致果实的酸度大。同时,相关性分析发现,避雨处理土壤微生物属水平上的变化趋势与梨的蔗糖含量和还原糖含量相关性最大,通过定向调控土壤微生物种群的变化,提高梨的品质。
李艳丽等[33]利用土壤覆盖管理方式研究不同土壤管理方式差异对梨园土壤微生物的影响,发现盖膜处理使土壤中的速效养分减少,而本研究中避雨栽培降低了土壤中速效磷、速效钾、有机质含量和含水率,两者研究结果一致。这可能是由于避雨处理梨树生长的要比露地处理的好,需要的土壤速效营养物质更多,最终导致果实品质更好。梨树生长需要土壤提供有效的养分和水分,在同等情况下,避雨处理防止了雨水进入土壤,减少了土壤含水率,就可能减少了土壤中可溶性养分的溶解与释放。本研究中相关性分析发现避雨处理土壤梨根际微生物的变化和土壤总氮、速效氮和pH显著正相关,而氮素是植物生长主要的营养元素。因此,可以通过调控梨园的氮素水平来调控土壤根际微生物。植物可以改变根际土壤微生物,根际土壤微生物也可以显著影响植物的生长、发育和对土源性病原体的抗性[34]。本研究发现避雨处理可以增加梨树根际土壤微生物的α多样性和丰富度,而微生物多样性和丰富度的增加可以提高根际微生态的稳定性和抗性。相关研究表明利用盖膜方式管理梨园可以增加梨树土壤微生物数量[33, 35],这与本研究结果一致。虽然,避雨处理虽然降低了土壤中K、P等有效养分和水分含量,但植物生长得更好,分泌更多的根系分泌物可以为根际微生物提供更多的营养物质,这可能是导致避雨处理梨树根际土壤微生物的多样性和丰富度要高于露地处理的原因。
4. 结论
生产实践结果证明避雨栽培对于梨的产量和品质都有提高作用。本研究表明,在山地种植的梨树,避雨栽培可以提高增加土壤微生物多样性和丰富度,增强梨树根际微生态系统的稳定性,增加梨树的抗逆性,从而提升梨树果实的品质。但是长期的盖棚对于土壤物质循环不利,不利于维持作物的长期生长需求。由于梨树避雨栽培更有利于植株生长,根系分泌物也较多的,这导致了土壤微生物数量和多样性有所增加,但是避雨处理不利于整个土壤物质循环,需要适度的补充肥料和水分。田间实际生产过程中应考虑长期的避雨处理带来的负面影响,通过适当的露地敞开时间可以改善土壤物质循环,有利于梨园生产的可持续和稳定性。
-
![]()
图 2 不同处理下碳降解的关键功能基因标准化相对信号强度
Figure 2. Normalized relative signal strength of key functional genes relating to C-degradation under different treatments
![]()
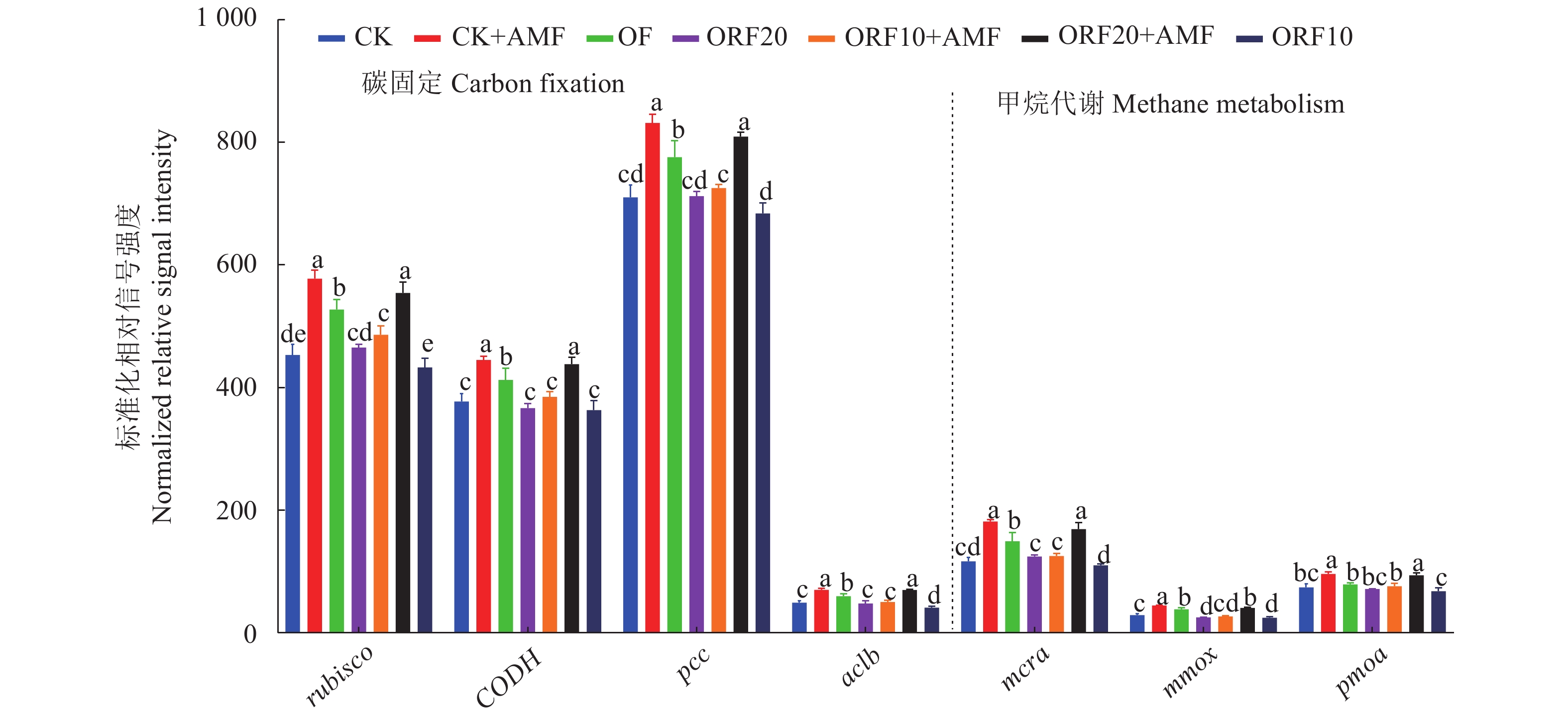
图 3 不同处理下碳固定与甲烷代谢的关键功能基因标准化相对信号强度
Figure 3. Normalized relative signal strength of key functional genes relating to C-fixation and
![]()
图 4 碳循环基因与理化性质的冗余分析
注:TN,总氮;AP,有效磷;TP,全磷;OM,有机质;TK,总钾;AK,速效钾;SR,土壤呼吸。
Figure 4. Redundant analysis on carbon cycle genes and physicochemical properties
Note: TN- total nitrogen; AP- available phosphorus; TP- total phosphorus; OM- organic matter; TK- total potassium; AK- available potassium; SR- soil respiration.
表 1 不同处理施肥方案
Table 1 Fertilization treatments
处理 Treatment 施肥用量 Fertilization dosage/(kg·hm-2) N P2O5 K2O 有机肥 Organic fertilizer CK,不施氮肥 0 150 225 OF,优化施肥 330 150 225 ORF10,有机肥氮替代10%化学氮肥 297 150 225 1650 ORF20,有机肥氮替代20%化学氮肥 264 150 225 3300 ORF10+AMF,有机肥氮替代10%化学氮肥+增施变形球囊霉菌(G. versiforme) 297 150 225 1650 ORF20+AMF,有机肥氮替代20%化学氮肥+增施变形球囊霉菌(G. versiforme) 264 150 225 3300 CK+AMF,不施氮肥+增施变形球囊霉菌(G. versiforme) 0 150 225  下载: 导出CSV
下载: 导出CSV
表 2 不同施肥下甜玉米产量与理化性质分析
Table 2 Yield and physicochemical properties of sweet corn under different treatments
处理 Treatment 产量 Yield/(kg·hm−2) 有机质含量 Organic content/(g·kg−1) 土壤呼吸 Soil respiration/(kg·hm−2·h−1) CK 7288.00±288.79 d 21.98±0.15 b 38.58±2.60 c OF 14246.20±469.13 b 22.06±0.60 b 54.64±5.68 b ORF10 14651.00±275.12 b 23.16±1.69 b 40.25±0.96 c ORF20 14925.00±855.37 b 24.03±2.06 ab 47.76±4.17 bc ORF10+AMF 15911.00±325.41 a 22.88±1.21 b 53.39±4.26 b ORF20+AMF 16250.00±630.72 a 25.97±2.57 a 74.66±10.02 a CK+AMF 9663.00±764.32 c 21.86±0.19 b 45.05±8.42 bc 显著性Significance P值 P Value AMF <0.001 <0.476 <0.001 有机无机配施(OCF) <0.001 <0.018 <0.001 AMF×ORF <0.191 <0.380 <0.029 注:同列数据后不同小写字母表示处理间差异显著性(P<0.05)。数据为均值 ± 标准误。
Note: Data with different lowercase letters on the same column indicate significant differences at P<0.05. Data presented as mean±standard error, n=3.
下载: 导出CSV
-
[1] BARDGETT R D, FREEMAN C, OSTLE N J. Microbial contributions to climate change through carbon cycle feedbacks [J]. The ISME Journal, 2008, 2(8): 805−814. DOI: 10.1038/ismej.2008.58
[2] ZHOU J Z, XUE K, XIE J P, et al. Microbial mediation of carbon-cycle feedbacks to climate warming [J]. Nature Climate Change, 2012, 2(2): 106−110. DOI: 10.1038/nclimate1331
[3] TRIVEDI P, ANDERSON I C, SINGH B K. Microbial modulators of soil carbon storage: Integrating genomic and metabolic knowledge for global prediction [J]. Trends in Microbiology, 2013, 21(12): 641−651. DOI: 10.1016/j.tim.2013.09.005
[4] 郑慧芬, 曾玉荣, 叶菁, 等. 农田土壤碳转化微生物及其功能的研究进展 [J]. 亚热带农业研究, 2018, 14(3):209−216. ZHENG H F, ZENG Y R, YE J, et al. Research advance on soil carbon conversion microorganisms and their functions in farmland ecosystems [J]. Subtropical Agriculture Research, 2018, 14(3): 209−216.(in Chinese)
[5] 王立, 贾文奇, 马放, 等. 菌根技术在环境修复领域中的应用及展望 [J]. 生态环境学报, 2010, 19(2):487−493. DOI: 10.3969/j.issn.1674-5906.2010.02.043 WANG L, JIA W Q, MA F, et al. Perspective of mycorrhizal technology application for environmental remediation [J]. Ecology and Environmental Sciences, 2010, 19(2): 487−493.(in Chinese) DOI: 10.3969/j.issn.1674-5906.2010.02.043
[6] 陆大雷, 刘小兵, 赵久然, 等. 甜玉米氮素吸收利用的基因型差异 [J]. 植物营养与肥料学报, 2008, 14(2):258−263. DOI: 10.3321/j.issn:1008-505X.2008.02.009 LU D L, LIU X B, ZHAO J R, et al. Genotypic differences in nitrogen uptake and utilization of sweet maize [J]. Plant Nutrition and Fertilizer Science, 2008, 14(2): 258−263.(in Chinese) DOI: 10.3321/j.issn:1008-505X.2008.02.009
[7] 张白鸽, 李强, 陈琼贤, 等. 广东甜玉米施肥指标体系研究 [J]. 广东农业科学, 2013, 40(20):67−70. DOI: 10.3969/j.issn.1004-874X.2013.20.022 ZHANG B G, LI Q, CHEN Q X, et al. Research on index system for sweet maize fertilization in Guangdong [J]. Guangdong Agricultural Sciences, 2013, 40(20): 67−70.(in Chinese) DOI: 10.3969/j.issn.1004-874X.2013.20.022
[8] 田善义, 王明伟, 成艳红, 等. 化肥和有机肥长期施用对红壤酶活性的影响 [J]. 生态学报, 2017, 37(15):4963−4972. TIAN S Y, WANG M W, CHENG Y H, et al. Long-term effects of chemical and organic amendments on red soil enzyme activities [J]. Acta Ecologica Sinica, 2017, 37(15): 4963−4972.(in Chinese)
[9] 郭良栋, 田春杰. 菌根真菌的碳氮循环功能研究进展 [J]. 微生物学通报, 2013, 40(1):158−171. GUO L D, TIAN C J. Progress of the function of mycorrhizal fungi in the cycle of carbon and nitrogen [J]. Microbiology China, 2013, 40(1): 158−171.(in Chinese)
[10] 金海如, 蒋湘艳, 夏婷婷. 不同有机物料及其菌根化对甜玉米产量与品质的协同影响 [J]. 中国土壤与肥料, 2019(6):196−203. JIN H R, JIANG X Y, XIA T T. Synergistic effect of different organic matters and mycorrhizal fungi on biomass and quality of sweet maize [J]. Soil and Fertilizer Sciences in China, 2019(6): 196−203.(in Chinese)
[11] 徐如玉, 左明雪, 袁银龙, 等. 增施摩西管柄囊霉对甜玉米氮肥增效及土壤丛枝菌根真菌多样性的影响 [J]. 福建农业学报, 2020, 35(4):379−391. XU R Y, ZUO M X, YUAN Y L, et al. Effects of Funneliformis mosseae application on nitrogen utilization by sweet corn and AM fungi diversity in soil [J]. Fujian Journal of Agricultural Sciences, 2020, 35(4): 379−391.(in Chinese)
[12] 徐丽娇, 姜雪莲, 郝志鹏, 等. 丛枝菌根通过调节碳磷代谢相关基因的表达增强植物对低磷胁迫的适应性 [J]. 植物生态学报, 2017, 41(8):815−825. DOI: 10.17521/cjpe.2017.0018 XU L J, JIANG X L, HAO Z P, et al. Arbuscular mycorrhiza improves plant adaptation to phosphorus deficiency through regulating the expression of genes relevant to carbon and phosphorus metabolism [J]. Chinese Journal of Plant Ecology, 2017, 41(8): 815−825.(in Chinese) DOI: 10.17521/cjpe.2017.0018
[13] 张弘. 相同碳氮比有机物料和生物炭对烤烟品质及土壤碳氮代谢的影响 [D]. 郑州: 河南农业大学, 2017. ZHANG H. Effect of the same C/N ratio organic material and boichar on flue-cured tobacco quality and soil carbon nitrogen metabolism [D]. Zhengzhou: Henan Agricultural University, 2017.(in Chinese)
[14] 梁晋刚, 辛龙涛, 栾颖, 等. 转cry1Ie基因抗虫玉米对根际微生物群落碳代谢的影响 [J]. 中国农业科技导报, 2019, 21(2):104−110. LIANG J G, XIN L T, LUAN Y, et al. Effect of Cry1Ie bt maize on carbon source metabolism of rhizosphere microorganisms [J]. Journal of Agricultural Science and Technology, 2019, 21(2): 104−110.(in Chinese)
[15] 路花, 张美俊, 冯美臣, 等. 氮肥减半配施有机肥对燕麦田土壤微生物群落功能多样性的影响 [J]. 生态学杂志, 2019, 38(12):3660−3666. LU H, ZHANG M J, FENG M C, et al. Effects of half-reduced nitrogen fertilization combined with organic fertilizer on functional diversity of soil microbial communities in oat field [J]. Chinese Journal of Ecology, 2019, 38(12): 3660−3666.(in Chinese)
[16] 孙丹萍. 丛枝菌根真菌扩繁技术研究 [J]. 河南林业科技, 2004, 24(2):12−13, 15. DOI: 10.3969/j.issn.1003-2630.2004.02.006 SUN D P. Study on the propagation technology of arbuscular mycorrhizal fungi [J]. Journal of Henan Forestry Science and Technology, 2004, 24(2): 12−13, 15.(in Chinese) DOI: 10.3969/j.issn.1003-2630.2004.02.006
[17] 毕银丽, 孙欢, 郭楠, 等. 不同基质和菌种组合对丛枝菌根真菌扩繁效果的影响 [J]. 应用与环境生物学报, 2017, 23(4):616−621. BI Y L, SUN H, GUO N, et al. Propagate-effects of different substrates and strain combinations on arbuscular mycorrhizal fungi [J]. Chinese Journal of Applied and Environmental Biology, 2017, 23(4): 616−621.(in Chinese)
[18] ZHAO Y P, LIN S, CHU L X, et al. Insight into structure dynamics of soil microbiota mediated by the richness of replanted Pseudostellaria heterophylla [J]. Scientific Reports, 2016, 6: 26175. DOI: 10.1038/srep26175
[19] 鲍士旦. 土壤农化分析 [M]. 北京: 中国农业出版社, 2000. [20] 鲁如坤. 土壤农业化学分析方法 [M]. 北京: 中国农业科技出版社, 2000. [21] STRALIS-PAVESE N, ABELL G C J, SESSITSCH A, et al. Analysis of methanotroph community composition using a pmoA -based microbial diagnostic microarray [J]. Nature Protocols, 2011, 6(5): 609. DOI: 10.1038/nprot.2010.191
[22] 吕鹏, 张吉旺, 刘伟, 等. 施氮量对超高产夏玉米产量及氮素吸收利用的影响 [J]. 植物营养与肥料学报, 2011, 17(5):852−860. LÜ P, ZHANG J W, LIU W, et al. Effects of nitrogen application on yield and nitrogen use efficiency of summer maize under super-high yield conditions [J]. Plant Nutrition and Fertilizer Science, 2011, 17(5): 852−860.(in Chinese)
[23] 段媛君, 王百田. 不同肥料与AM真菌配施对沙打旺品质的影响 [J]. 西北农林科技大学学报(自然科学版), 2019, 47(5):118−124. DUAN Y J, WANG B T. Effect of different fertilizers and AM fungi on quality of Astragalus adsurgens Pall [J]. Journal of Northwest A & F University (Natural Science Edition), 2019, 47(5): 118−124.(in Chinese)
[24] 周世品. 有机无机肥配施和丛枝菌根化育苗对西瓜产量品质的影响 [D]. 南京: 南京农业大学, 2017. ZHOU S P. Effects of combined organic and inorganic fertilizer and arbuscular mycorrhizal colonization on watermelon yield and quality [D]. Nanjing: Nanjing Agricultural University, 2017.(in Chinese)
[25] 张前兵. 干旱区不同管理措施下绿洲棉田土壤呼吸及碳平衡研究 [D]. 石河子: 石河子大学, 2013. ZHANG Q B. Studies on soil respiration and carbon balance under different management practices in cotton field of oasis in arid region[D]. Shihezi: Shihezi University, 2013.(in Chinese)
[26] 贺美,王迎春,王立刚,等.不同耕作措施对黑土碳排放和活性碳库的影响[J].土壤通报,2016, 47(5):1195-1202. HE M, WANG Y C, WANG L G, et al. Effect of different tillage managements oncarbon dioxide emission and content of activated carbon in black soil [J]. Chinese Journal of Soil Science, 2016, 47(5): 1195-1202.(in Chinese)
[27] HODGE A, FITTER A H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling [J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(31): 13754−13759. DOI: 10.1073/pnas.1005874107
[28] CHENG L, BOOKER F L, TU C, et al. Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2 [J]. Science, 2012, 337(6098): 1084−1087. DOI: 10.1126/science.1224304
[29] RUI J P, LI J B, WANG S P, et al. Responses of bacterial communities to simulated climate changes in alpine meadow soil of the Qinghai-Tibet plateau [J]. Applied and Environmental Microbiology, 2015, 81(17): 6070−6077. DOI: 10.1128/AEM.00557-15
[30] TABITA F R. Microbial ribulose1,5-bisphosphate carboxylase/oxygenase: A different perspective [J]. Photosynthesis Research, 1999, 60(1): 1−28. DOI: 10.1023/A:1006211417981
[31] SIEGENTHALER U, SARMIENTO J L. Atmospheric carbon dioxide and the ocean [J]. Nature, 1993, 365(6442): 119−125. DOI: 10.1038/365119a0
[32] 刘洋荧, 王尚, 厉舒祯, 等. 基于功能基因的微生物碳循环分子生态学研究进展 [J]. 微生物学通报, 2017, 44(7):1676−1689. LIU Y Y, WANG S, LI S Z, et al. Advances in molecular ecology on microbial functional genes of carbon cycle [J]. Microbiology China, 2017, 44(7): 1676−1689.(in Chinese)
[33] GIFFORD R M. The global carbon cycle: A viewpoint on the missing sink [J]. Functional Plant Biology, 1994, 21(1): 1. DOI: 10.1071/PP9940001
[34] VAN GROENIGEN K J, OSENBERG C W, HUNGATE B A. Increased soil emissions of potent greenhouse gases under increased atmospheric CO2 [J]. Nature, 2011, 475(7355): 214−216. DOI: 10.1038/nature10176
[35] XUE K, YUAN M M, SHI Z J, et al. Tundra soil carbon is vulnerable to rapid microbial decomposition under climate warming [J]. Nature Climate Change, 2016, 6(6): 595−600. DOI: 10.1038/nclimate2940
[36] PENDALL E, BRIDGHAM S D, HANSON P J, et al. Below-ground process responses to elevated CO2 and temperature: A discussion of observations, measurement methods, and models [J]. New Phytologist, 2004, 162(2): 311−322. DOI: 10.1111/j.1469-8137.2004.01053.x
[37] FORSTER P, RAMASWAMY V, ARTAXO P, et al. Changes in atmospheric constituents and in radiative forcing [M] the 4th Assessment Report of the IPCC WG1: The Physical Science Basis. DLR, 2007.
[38] HE Z L, XU M Y, DENG Y, et al. Metagenomic analysis reveals a marked divergence in the structure of belowground microbial communities at elevated CO2 [J]. Ecology Letters, 2010, 13(5): 564−575. DOI: 10.1111/j.1461-0248.2010.01453.x
[39] YERGEAU E, KANG S, HE Z L, et al. Functional microarray analysis of nitrogen and carbon cycling genes across an Antarctic latitudinal transect [J]. The ISME Journal, 2007, 1(2): 163−179. DOI: 10.1038/ismej.2007.24
[40] REEVE J R, SCHADT C W, CARPENTER-BOGGS L, et al. Effects of soil type and farm management on soil ecological functional genes and microbial activities [J]. The ISME Journal, 2010, 4(9): 1099−1107. DOI: 10.1038/ismej.2010.42
[41] LIU Z F, LIU G H, FU B J, et al. Relationship between plant species diversity and soil microbial functional diversity along a longitudinal gradient in temperate grasslands of Hulunbeir, Inner Mongolia, China [J]. Ecological Research, 2008, 23(3): 511−518. DOI: 10.1007/s11284-007-0405-9
[42] WARDLE D A. Ecological linkages between aboveground and belowground biota [J]. Science, 2004, 304(5677): 1629−1633. DOI: 10.1126/science.1094875
[43] ZHANG Y G, CONG J, LU H, et al. An integrated study to analyze soil microbial community structure and metabolic potential in two forest types [J]. PLoS One, 2014, 9(4): e93773. DOI: 10.1371/journal.pone.0093773
[44] ROUSK J, BROOKES P C, BAATH E. The microbial PLFA composition as affected by pH in an arable soil [J]. Soil Biology & Biochemistry, 2010, 42(3): 516−520.
[45] BASTIDA F, MORENO J L, HERNÁNDEZ T, et al. Microbiological activity in a soil 15 years after its devegetation [J]. Soil Biology and Biochemistry, 2006, 38(8): 2503−2507. DOI: 10.1016/j.soilbio.2006.02.022
[46] LI Y Q, XU M, ZOU X M, et al. Soil CO2 efflux and fungal and bacterial biomass in a plantation and a secondary forest in wet tropics in Puerto Rico [J]. Plant and Soil, 2005, 268(1): 151−160. DOI: 10.1007/s11104-004-0234-3
-
期刊类型引用(6)
1. 李文帅,段玉春,范文艳,韩文革,吕清晗,吴皓菘,李明睿,李宝成,姜述君. 节杆菌和腐熟鸡粪混施对Atrazine污染土壤特性影响. 黑龙江八一农垦大学学报. 2024(03): 18-24 .  百度学术
百度学术
2. 郑一玲,范栩妙,钟哲伦,姚光伟,林晨,顾佳艳,靳泽文,平立凤,单胜道. 生物质炭-沼液配施条件下旱地红壤碳氮循环功能基因丰度主控因子与耦合关系. 农业资源与环境学报. 2024(04): 835-845 . 百度学术
3. 马啸驰,韩烽,白亚涛,吴双,吴景贵,马艳. 有机物料在果园生态系统中的应用及其环境效应:研究现状与展望. 中国生态农业学报(中英文). 2023(08): 1240-1255 . 百度学术
4. 乔志伟,刘超,王红. 一株硝基还原假单胞菌的溶磷特性及对碳循环相关基因的影响. 江苏农业学报. 2023(05): 1151-1158 . 百度学术
5. 马慧霞,张桥,陈会巧,孙丽丽,周昌敏,顾文杰,卢钰升,徐培智,解开治. 长期有机培肥对南方红壤区稻田土壤碳循环功能基因的影响. 南方农业学报. 2023(05): 1405-1416 . 百度学术
6. 顾美英,唐光木,张云舒,黄建,张志东,张丽娟,朱静,唐琦勇,楚敏,徐万里. 有机肥与生物炭对新疆盐碱沙化土壤微生物群落特征的影响. 生态环境学报. 2023(08): 1392-1404 . 百度学术
其他类型引用(16)







计量
- 文章访问数: 1254
- HTML全文浏览量: 315
- PDF下载量: 54
- 被引次数: 22
