

Genetics of Lagerstroemia indica × L. limii Phenotype
-
摘要:目的
探讨福建紫薇(Lagerstroemia limii)改良紫薇乔木性状的潜力。
方法以福建紫薇与灌木型紫薇品种‘千层绯雪’’为亲本构建正、反交群体,采用相关性分析、主基因+多基因混合遗传模型对一年生杂交后代的株高、地径、一级分枝点高度、一级分枝数等7个性状进行分析。
结果(1)杂交后代的株高在1.50~24.5 cm,地径0.30~4.44 mm,一级分枝点高度0.50~6.20 cm,一级分枝数0~4个,多数性状基本呈正态分布,后代中存在潜在的乔木型种质。(2)7个性状的变异系数范围在31.72%~240.42%,变异程度最大的是千层绯雪×福建紫薇群体的一级分枝数,最小的是该群体的一级分枝点高度。福建紫薇ב千层绯雪’群体的一级分枝点高度、开张角度、侧枝GSA的变异系数比‘千层绯雪’×福建紫薇群体更大,且福建紫薇ב千层绯雪’群体更倾向表现出植株高、地径粗、分枝数量多,说明福建紫薇作为母本时杂交后代有更丰富的株型变异。(3)相关性分析表明,株高与地径、一级分枝数与开张角度、开张角度与侧枝GSA呈极显著正相关。(4)株高、地径、开张角的最适模型为2MG-AD,侧枝GSA、一级分枝点高度的最适模型为2MG-EA。
结论福建紫薇是培育生长量大、基部不分枝、主干明显的紫薇品种的优良材料,为乔木型紫薇的培育提供依据。
Abstract:ObjectiveGenetics on the phenotype of Lagerstroemia limii was studied to breed improved hybrids with 'Qianceng Feixue'.
MethodPopulations of hybrids between the tree-type L. limii (FJ) and the shrub-type 'Qianceng Feixue'(QC) were generated for the study. Seven phenotype traits including the height, diameter, and length of 1st branch of one-year-old plants were measured for correlation analysis among them. Major gene+polygene inheritance models were tested for best fitting in predicting hybrid phenotype.
Result(1) The plant height of the hybrid progenies ranged 1.50–24.5 cm, the trunk diameter 0.30–4.44 mm, the 1st branch length 0.50–6.20 cm, and the branch number 0–4. These traits were basically in a normal distribution pattern among the hybrids with potential arborescent germplasms. (2) The coefficients of variation on the 7 traits were 31.72%–240.42% with the greatest on the number of branches and the least on the height of the first branch in the QC×FJ population. The coefficients of variation of primary branching point height, opening angle, and lateral branching GSA were larger in the FJ ×QC population than in the QC × FJ population, and the FJ × QC group tended to show higher plant height, thicker ground diameter, and more branches, indicating that the hybrid progeny of L. limii as the mother plant had richer strain variation. (3) The plant height significantly correlated with the trunk diameter, the number with the angle of the branches, and the angle with the GSA of the branches. (4) The best fit model for the plant height, trunk diameter, and branching angle was 2MG-AD, and that for the branch GSA and 1st branch length 2MG-EA.
ConclusionL. limii (FJ) appeared to be a plausible candidate for breeding hybrids with L. indica (QC) that could inherit the desirable phenotypes of growing vigorously, being free of branches on base of trunk, and showing the main trunk prominently.
-
Keywords:
- Lagerstroemia /
- distant hybridization /
- phenotype traits /
- genetic analysis
-
0. 引言
【研究意义】随着国民经济的发展和人民生活水平的不断提高,人们不再满足于填饱肚子的需求,进而对食用稻米的品质提出了更高的要求[1]。粳稻与籼稻相比,具有口感好、黏度高、营养价值丰富的特点,因而越来越受到人们的青睐[2−3],近十年来,我国粳稻播种面积和单产均有明显的提升,南方稻区也有籼改粳的发展趋势[4−5],目前,我国粳稻种植面积已达990. 43 万hm2 ,总产量7 064. 85万t,比2010年增加约
1200 万t[6]。当前中国稻米市场对粳米的需求持续增长,我国城乡居民尤其是长江中下游地区居民的稻米消费出现由籼米向粳米转变的趋势[7]。2019年我国粳稻的人均消费量为55.4 kg,比2000年人均消费增加17.6 kg[8]。因此,粳稻生产在我国水稻生产中占据越来越重要的地位。我国粳稻资源丰富,分布广泛,31个省(市、区)均有分布,粳稻资源有17858 份,入库保存的有15791 份[9]。【前人研究进展】杨忠义等[10]对中国地方粳稻资源特征特性评价及多样性分布研究结果表明中国粳稻资源特征特性丰富多样,蕴藏着丰富的优异基因;沈新平等[11]对368份太湖地区晚粳稻地方种的分析结果表明,种质间的性状均有极显著差异;陈小龙等[12]对宁夏60份粳稻种质资源材料的16个表型性状进行遗传多样性分析结果表明种质的表型性状间存在极显著的遗传差异;陈丽等[13]对60份来源不同的粳稻表型性状多样性及综合评价表明18个表型性状的变异系数在1.10%~63.40%,筛选获得综合得分最高的前10 名种质。【本研究切入点】作为全国水稻育种强省,福建省在籼稻生产全面发展的基础上最大程度地发展粳稻生产,培育出适应福建省乃至全国推广应用的粳稻新品种,对保障我国粮食安全和提高稻作生产效益具有重要意义。【拟解决的关键问题】本研究利用变异系数、遗传多样性指数、相关分析方法、聚类分析及主成分分析等统计学方法,对新引进的140份粳稻种质资源进行表型性状多样性分析及综合评价,以明确粳稻种质的表型多样性及其综合表型性状评价指标,为高效利用粳稻资源、拓宽遗传基础、创制粳稻新种质提供参考依据。1. 材料与方法
1.1 试验材料
试验材料包括140份粳稻资源(表1),从国家粮食作物种质资源中期库引进。
表 1 供试材料的亚种类型及来源Table 1. Subspecies and source of germplasms序号
Number品种名
Germplasm name亚种类型
Subspecies type来源地
Origin序号
Number品种名
Germplasm name亚种类型
Subspecies type来源地
Origin1 农科843 粳稻 宁夏 71 通系12 粳稻 吉林 2 丹梗8 粳稻 辽宁 72 吉粳81 粳稻 吉林 3 辽星1 粳稻 辽宁 73 垦优109 粳稻 河北 4 沈农315 粳稻 辽宁 74 垦育38 粳稻 河北 5 铁粳4号 粳稻 辽宁 75 垦稻09-1 粳稻 河北 6 抚粳8号 粳稻 辽宁 76 垦糯8 粳稻 河北 7 旱糯303 粳稻 辽宁 77 中作0516 粳稻 北京 8 盐粳48 粳稻 辽宁 78 中作0332 粳稻 北京 9 福粳 2103 粳稻 辽宁 79 千重浪1号 粳稻 辽宁 10 新宾1号 粳稻 辽宁 80 中丹4号 粳稻 辽宁 11 平壤一号 粳稻 朝鲜 81 中辽 9052 粳稻 辽宁 12 超产1号 粳稻 吉林 82 花粳15 粳稻 辽宁 13 松粳12 粳稻 黑龙江 83 盐丰47 粳稻 辽宁 14 梦粳26号 粳稻 云南 84 吉粳83 粳稻 吉林 15 定育1号 粳稻 朝鲜 85 松粳9号 粳稻 黑龙江 16 银龟 粳稻 朝鲜 86 松粳11 粳稻 黑龙江 17 梦结 粳稻 日本 87 松粳13 粳稻 黑龙江 18 奥村紫 粳稻 日本 88 龙稻2 粳稻 黑龙江 19 脂光 粳稻 日本 89 龙稻3 粳稻 黑龙江 20 银宝 粳稻 日本 90 悬桥 粳稻 日本 21 中花15号 粳稻 北京 91 BG1 粳稻 韩国 22 郑旱2号 粳稻 河南 92 雪香稻 粳稻 韩国 23 郑旱6号 粳稻 河南 93 花114 粳稻 宁夏 24 毕粳37 粳稻 贵州 94 7-218 粳稻 宁夏 25 华吹雪 粳稻 日本 95 京宁3号 粳稻 宁夏 26 美山锦 粳稻 日本 96 京宁6号 粳稻 宁夏 27 秋田小町 粳稻 日本 97 新子7 粳稻 河北 28 岩手传手 粳稻 日本 98 新子10 粳稻 河北 29 里歌 粳稻 日本 99 丰民 2102 粳稻 辽宁 30 接冠 粳稻 日本 100 丹137 粳稻 辽宁 31 一目惚 粳稻 日本 101 丹粳9号 粳稻 辽宁 32 桂桥 粳稻 日本 102 丹粳11 粳稻 辽宁 33 驹舞 粳稻 日本 103 大关稻 粳稻 韩国 34 奥羽387 粳稻 日本 104 南一稻 粳稻 韩国 35 银龟12号 粳稻 朝鲜 105 右流间糯 粳稻 日本 36 登系52号 粳稻 朝鲜 106 宝石稻 粳稻 韩国 37 雄基3号 粳稻 朝鲜 107 中大菲 粳稻 广西 38 咸南23号 粳稻 朝鲜 108 七矮占 粳稻 广西 39 境城8号 粳稻 朝鲜 109 华粳42 粳稻 贵州 40 Q568 粳稻 贵州 110 农虎禾 粳稻 贵州 41 水原300粒 粳稻 河北 111 华想 粳稻 日本 42 中楼一号 粳稻 山西 112 安山稻 粳稻 韩国 43 兴图 粳稻 吉林 113 黑香稻 粳稻 韩国 44 京虎B 粳稻 安徽 114 云粳33号 粳稻 云南 45 G珍汕 9713 粳稻 四川 115 风稻11号 粳稻 云南 46 粳 7623 粳稻 上海 116 风稻14号 粳稻 云南 47 宁E21 粳稻 江苏 117 风稻15号 粳稻 云南 48 76-1 粳稻 辽宁 118 风稻17号 粳稻 云南 49 中花8号 粳稻 北京 119 绿苑糯稻 粳稻 韩国 50 晋稻1号 粳稻 山西 120 早生统一 粳稻 韩国 51 辽粳287 粳稻 辽宁 121 斗立 粳稻 韩国 52 当育5号 粳稻 安徽 122 珍珠1号 粳稻 韩国 53 日引A8 粳稻 日本 123 里的歌 粳稻 日本 54 日引A9 粳稻 日本 124 神锦 粳稻 日本 55 西珍稻 粳稻 韩国 125 一见钟情选 粳稻 日本 56 珍富36号 粳稻 韩国 126 云冷22号 粳稻 云南 57 高山稻 粳稻 韩国 127 云冷25号 粳稻 云南 58 雪香糯稻 粳稻 韩国 128 合系15 粳稻 云南 59 早安稻 粳稻 韩国 129 05-1876 粳稻 北京 60 西平稻 粳稻 韩国 130 06- 2391 粳稻 北京 61 青湖稻 粳稻 韩国 131 水原55号 粳稻 韩国 62 风味稻 粳稻 韩国 132 里里372 粳稻 韩国 63 云光稻 粳稻 韩国 133 桂花黄 粳稻 江苏 64 新尚州稻 粳稻 韩国 134 晋稻1号 粳稻 山西 65 咸州3号 粳稻 朝鲜 135 辽粳287 粳稻 辽宁 66 山光 粳稻 日本 136 福锦 粳稻 日本 67 2004-460 粳稻 宁夏 137 田锦糯 粳稻 日本 68 节12 粳稻 宁夏 138 藤834 粳稻 日本 69 2008XZ-89 粳稻 宁夏 139 藤842 粳稻 日本 70 宁大 12596 粳稻 宁夏 140 巴利拉 粳稻 意大利 1.2 田间扩繁与表型性状鉴定
供试材料于2017—2018年在福建省将乐县古镛镇张公村(东经117°27′11.282″,北纬26°44′51.191″)进行扩繁收种与鉴定评价,每份材料种植1个小区,每小区种植15行,每行7株,共105株,株行距20 cm×20 cm。肥水管理与当地大田生产相同。参照《水稻种质资源描述规范和数据标准》[14]调查抽穗期、全生育期、株高、穗长、穗粒数、有效穗数、结实率、千粒重、谷粒长度、谷粒宽度、谷粒形状等11个项目。
1.3 数据统计与分析
使用 Excel 2016 整理数据,利用 SPSS 25.0 对表型性状进行数据统计、遗传多样性指数、相关性状分析和主成分分析,使用Origin 2021 进行聚类分析及绘图。
2. 结果与分析
2.1 140份粳稻资源表型性状分布特征
140份粳稻资源10个性状分布特征见表2,所有资源均能正常抽穗,抽穗期在2022-07-06—2022-09-01,抽穗期主要集中在7、8月份,占99.3%,9月份抽穗仅有1份;全生育期变幅在86~128 d,主要集中在86~110 d,占87.8%;株高变幅在68.18~144.02 cm,主要集中在中矮和中两种类别,占85.7%;穗长变幅在14.36~28.67 cm,只有短和中两种类别;有效穗数变幅在5.80~21.80个,主要集中在少和中两种类别,占97.8%;穗粒数变幅在45.40~317.40,以中等粒型为主,占63.6%,少和多两种类别共占32.9%;结实率变幅在41.51%~91.95%,中等类型占50.7%,低和高两种类别共占47.1%;千粒重变幅在17.23~34.61 g,以中等类别为主,占92.9%;谷粒长度变幅在6.26~9.28 mm,以中等类别为主,占87.1%;谷粒宽度变幅在2.29~4.28 mm,以中等类别为主,占91.4%;谷粒形状以椭圆形为主,占82.9%。10个性状各类别的分布情况表明140份粳稻资源以中间类别为主,同时存在少量极端类别,表型性状遗传变异丰富。
表 2 10个数量性状频率分布Table 2. Frequency distribution of 10 quantitative traits项目
Project各类别分布情况
Distribution of all kinds of categories全生育期
Growth duration/d≤ 100 (100,110] (110,120] (120, 130] >130 比例
Proportion/%32.1 55.7 8.6 3.6 0 株高
Plant height/cm矮(≤70.0) 中矮(70.0,90.0] 中(90.0,110.0] 中高(110.0,130.0] 高(>130.0) 比例
Proportion/%1.4 33.6 52.1 10.7 2.1 穗长
Panicle length/cm极短(≤10.0) 短(10.0,20.0] 中(20.0,30.0] 长(30.0,40.0] 极长(>40) 比例
Proportion/%0 60.0 40.0 0 0 有效穗数
Panicles per plant极少(≤5) 少(5,10] 中(10,20] 多(>20) 比例
Proportion/%0 20.7 77.1 2.1 穗粒数
Spikelets per panicle极少(≤60) 少(60,100] 中(100,200] 多(200,300] 极多(>300) 比例
Proportion/%2.1 17.9 63.6 15.0 1.4 结实率
Seed setting rate/%不结实(0) 低(≤65.0) 中(65.0,80.0] 高(80.0,90.0] 极高(>90) 比例
Proportion/%0 20.7 50.7 26.4 2.1 千粒重 1000 -grain weight/g极低(≤10.0) 低(10.0,20.0] 中(20.0,30.0] 高(30.0,40.0] 极高(>40) 比例
Proportion/%0 5.7 92.9 1.4 0 谷粒长度
Grain length/mm极短(≤4.0) 短(4.0,6.0] 中(6.0,8.0] 长(8.0,10.0] 极长(>10) 比例
Proportion/%0 0 87.1 12.9 0 谷粒宽度
Grain width/mm极窄(≤1.5) 窄(1.5,2.5] 中(2.5,3.5] 宽(3.5,4.5] 极宽(>4.5) 比例
Proportion/%0 1.4 91.4 7.9 0 谷粒形状
Grain shape短圆形(≤1.80) 阔卵形(1.80,2.20] 椭圆形(2.20,3.00] 中长形(3.00,3.30] 细长形(>3.30) 比例
Proportion/%0 15.0 82.9 2.1 0 谷粒形状的数值是长宽比。
The value of grain shape is length-width ratio.2.2 140份粳稻资源表型性状的遗传变异和多样性分析
对10个表型性状进行统计分析(表3)结果表明,10个性状均存在不同程度的变异,变异幅度6.87%~36.86%,其中穗粒数的变异系数最大(36.86%),极差达272粒,谷粒长度变异系数最小(6.87%)。除全生育期、谷粒长度、谷粒宽度和谷粒形状外的其他6个性状的变异系数均大于10%。说明这批新引进的粳稻资源表型性状遗传变异丰富,可为粳稻育种和基础研究提供新种质。
表 3 10个数量性状的遗传变异性Table 3. Genetic variations on 10 quantifiable traits性状
Traits平均值
Mean标准差
SD极小值
Min极大值
Max极差
Range变异系数
CV/%全生育期
Growth duration/d102.81 8.05 86.00 128.00 42.00 7.83 株高
Plant height/cm96.21 13.08 68.18 144.02 75.84 13.60 穗长
Panicle length/cm19.94 2.72 14.36 28.67 14.31 13.65 有效穗数
Panicles per plant12.59 3.01 5.80 21.80 16.00 23.90 结实率
Seed setting rate/%72.79 10.74 41.51 91.95 50.44 14.75 穗粒数
Spikelets per panicle144.05 53.10 45.40 317.40 272.00 36.86 千粒重 1000 -grain weight/g24.19 2.45 17.23 34.61 17.38 10.14 谷粒长度
Grain length/mm7.52 0.52 6.26 9.28 3.02 6.87 谷粒宽度
Grain width/mm3.19 0.27 2.29 4.28 1.99 8.33 谷粒形状
Grain shape2.37 0.21 2.01 3.29 1.28 9.03 对10个表型性状进行遗传多样性分析(图1)。从图1可以看出,10个性状遗传多样性指数变化范围1.87~2.07,其中全生育期(1.95)、株高(1.96)、穗长(2.03)、有效穗数(2.07)、结实率(1.96)、穗粒数(1.98)、千粒重(2.02)、谷粒长(2.00)、谷粒宽(1.98)、谷粒形状(1.87),有效穗数的遗传多样性指数最高,谷粒形状的遗传多样性指数最低,极差值为0.20,总体上,这10个性状遗传多样性指数差异不大。
2.3 140份粳稻资源表型性状的相关性分析
对140份粳稻资源10个表型性状进行相关性分析(表4),结果表明,全生育期与株高、穗长和穗粒数呈极显著正相关,与结实率呈极显著负相关;株高与穗长、穗粒数和谷粒形状呈极显著正相关,与结实率和谷粒宽度呈显著负相关;穗长与穗粒数和谷粒形状极显著正相关,与谷粒宽度呈极显著负相关;有效穗数与穗粒数呈极显著负相关,与谷粒宽度呈显著负相关;结实率与千粒重和谷粒宽度呈极显著正相关,与穗粒数呈极显著负相关;穗粒数与谷粒形状呈显著正相关,穗粒数与千粒重呈极显著负相关;千粒重与谷粒长度和谷粒宽度呈极显著正相关,谷粒长度与谷粒宽度和谷粒形状呈极显著正相关,谷粒宽度与谷粒形状呈极显著负相关。
表 4 个数量性状相关性分析Table 4. Correlations among 10 quantifiable traits性状
Traits全生育期
Growth
duration株高
Plant
height穗长
Panicle
length有效穗数
Panicles
per plant结实率
Seed setting
rate穗粒数
Spikelets per
panicle千粒重 1000 -grain
weight谷粒长度
Grain
length谷粒宽度
Grain
width谷粒形状
Grain
shape全生育期
Growth duration1 株高
Plant height0.410** 1 穗长
Panicle length0.333** 0.710** 1 有效穗数
Panicles per plant−0.123 −0.038 −0.130 1 结实率
Seed setting rate−0.235** −0.172* −0.092 0.033 1 穗粒数
Spikelets per panicle0.500** 0.447** 0.458** −0.503** −0.396** 1 千粒重 1000 -grain weight−0.091 −0.036 −0.003 −0.062 0.225** −0.338** 1 谷粒长度
Grain length−0.092 0.081 0.146 −0.201* 0.224** −0.140 0.496** 1 谷粒宽度
Grain width−0.157 −0.172* −0.269** −0.100 0.080 −0.306** 0.538** 0.395** 1 谷粒形状
Grain shape0.081 0.253** 0.402** −0.062 0.119 0.195* −0.127 0.418** −0.661** 1 *表示差异达到显著水平(P<0.05),**表示差异达到极显著水平(P<0.01)。
* indicates significant difference at P<0.05; ** indicates extremely significant difference at P<0.01.2.4 140份粳稻资源10个表型性状的聚类分析
利用Origin 2021对原始数据进行聚类分析,构建140份粳稻资源的聚类图(图2),共聚为5类,各类群粳稻资源10个表型性状的平均值(表5),第Ⅰ类包括86份种质,占参试水稻资源的61.43%,这类群资源的生育期,株高和穗长在5个类型中最小;这类群的平均有效穗数在5个类型中最大,本类群具有早熟,株高较矮,穗长较短和多穗等特点。第Ⅱ类包括3份种质,分别是中楼1号、兴图、BG1,这类群资源的结实率、千粒重、谷粒长度和谷粒宽度在5个类型中最大,穗粒数在5个类型中最小;本类群具有大穗、大粒和高结实率等特点。第Ⅲ类包括20份种质,占参试水稻资源的14.29%,这类群资源的有效穗数在5个类型中最少;其他10个性状在5个类型中居中,本类群具有少穗等特点。第Ⅳ类包括29份种质,占参试水稻资源的20.71%,这类群资源的11个性状在5个类型中居中,本类群表型性状没有极端类型。第Ⅴ类包括3份种质,分别是中大菲、云冷22号,这类群资源的生育期、株高、穗长、穗粒数和谷粒形状在5个类型中最大,结实率、千粒重、谷粒长度和谷粒宽度在5个类型中最小;本类群具有生育期偏长,植株高大,穗大,粒多,小粒等特点。
表 5 5个类型的表型性状差异分析Table 5. Different phenotypic traits in 5 taxa类型
Type数量
Number全生育期
Growth
duration/d株高
Plant
height/cm穗长
Panicle
length/cm有效穗数
Panicles
per plant结实率
Seed setting
rate/%穗粒数
Spikelets per
panicle千粒重 1000 -grain
weight/g谷粒长度
Grain
length/mm谷粒宽度
Grain
width/mm谷粒形状
Grain
shapeⅠ 86 99.79±6.4 90.23±7.69 18.98±1.97 13.51±2.39 75.35±8.98 113.57±23.40 24.5±2.35 7.5±0.42 3.23±0.24 2.34±0.20 Ⅱ 3 100.67±4.04 117.12±10.21 22.01±1.20 12.67±2.50 80.73±5.27 64.33±18.18 27.52±6.14 8.2±0.73 3.46±0.71 2.4±0.30 Ⅲ 20 107.1±7.62 101.55±10.83 21.38±2.78 9.63±2.21 67.99±13.20 230.02±16.59 23.5±2.21 7.61±0.61 3.14±0.24 2.43±0.18 Ⅳ 29 107.86±8.28 105.09±13.56 20.97±2.96 11.93±3.81 68.82±10.73 171.52±20.26 23.51±2.11 7.46±0.65 3.15±0.22 2.37±0.23 Ⅴ 2 119.5±2.12 140.39±5.13 28.39±0.39 12.1±0.71 56.63±19.58 316.2±1.70 22.14±1.04 6.96±0.04 2.52±0.34 2.79±0.38 从图2和表5可以看出,5个类群粳稻资源的表型性状有明显差异,且每个类群中粳稻资源的数量不同,说明供试140份粳稻资源遗传背景较为复杂,具有丰富的遗传多样性。
2.5 140份粳稻资源10个表现性状的主成分分析
对140份粳稻资源表现性状的主成分分析(表6),共提取了4个主成分,累积贡献率为77.42%,这4个主成分可以代表140份粳稻资源10个表型性状的主要遗传信息。主成分1的特征值和贡献率分别是3.065和30.646%,主要包含株高和穗长2个作用因子;主成分2的特征值和贡献率分别是1.986和19.859%,主要包含千粒重和谷粒长度2个作用因子;主成分3的特征值和贡献率分别是
1.5827 和15.816%,谷粒宽度特征值的绝对值为0.777,谷粒宽度和谷粒形状是该主成分的主要作用因子;主成分4的特征值和贡献率分别是1.110和11.097%,有效穗数特征值为0.940远大于其他表型值;说明有效穗数为该主成分主要作用因子。分析各因子中主要指标的比重,可用株高、穗长、千粒重、谷粒长度、谷粒宽度、谷粒形状和有效穗数等指标来概括10个性状的大部分信息。表 6 10个表型性状的主成分分析Table 6. Principal components of 10 phenotypes性状
Trait主成分
Component1 2 3 4 全生育期 Growth duration 0.666 −0.190 −0.081 −0.186 株高 Plant height 0.881 0.066 0.141 0.049 穗长 Panicle length 0.797 0.140 0.332 −0.050 有效穗数 Panicles per plant −0.014 −0.150 0.000 0.940 结实率 Seed setting rate −0.366 0.465 0.300 0.187 穗粒数 Spikelets per panicle 0.557 −0.349 0.120 −0.652 千粒重 1000 -grain weight0.023 0.792 −0.321 0.080 谷粒长度 Grain length 0.044 0.864 0.186 −0.176 谷粒宽度 Grain width −0.136 0.544 −0.777 −0.052 谷粒形状 Grain shape 0.183 0.172 0.924 −0.086 特征值 Eigenvalue 3.065 1.986 1.582 1.11 贡献率 Contribution/% 30.646 19.859 15.816 11.097 累计贡献率 Cumulative contribution/% 30.646 50.505 66.321 77.418 2.6 140份粳稻资源表型性状综合评价和筛选
根据主成分分析结果,将140份稻种资源10个表型性状的标准化值代入4个主成分中,求得各种质的4个主成分得分。根据各主成分的累积贡献率(77.418%)得到前4个主成分的权重(
0.3959 、0.2565 、0.2043 、0.1433 ),通过权重计算得到140份稻种资源的综合评价D值,进而对各种质进行综合评价,D 值越高,表型性状的综合表现越好。综合排名前10的资源是当育5号、奥村学、中大菲、中花15号、云冷22号、BG1、农虎禾、毕梗42、兴图、G珍汕9713 。3. 讨论与结论
3.1 粳稻资源表型性状遗传多样性
开展水稻遗传多样性研究是水稻种质资源评价和保护的必要工作,也可以为育种家选育优良品种所需亲本提供有效依据。利用形态或表型性状研究水稻遗传多样性仍然是最简单、经济而直接的有效方法,尤其当研究材料群体较大时,通过观察表型性状能在较短的时间内对供试材料的遗传多样性水平有个初步认识,所以表型性状被广泛利用于亲本选择[15−17]。本研究对新引进的140份粳稻资源表型性状进行遗传变异和多样性分析,结果表明:表型性状的各类别分布以中间类型为主,也存在少量极端类型,有效穗>20的资源有3份,穗粒数>300的资源有2份,结实率>90%的资源有3份,千粒重在30~40 g的资源有2份,谷粒长度在8.0~10.0 mm的资源有18份,谷粒宽度在3.5~4.5 mm的资源有11份,筛选出3份细长型和21份阔卵型的粳稻资源。一般来说表型性状中越容易含有极端的变异类型,意味着可供筛选材料越多,培育出具有优异性状的品种机会越大。在本研究中,变异系数和遗传多样性指数在性状表达上没有一致性,如穗粒数变异系数最大但遗传多样性指数并不是最高,有效穗数遗传多样性指数最高但变异系数不是最高,其原因是它们反映遗传变异的内涵不同,变异系数反映的是某一性状变异范围,而遗传多样性指数则是指某一性状的不同表现等级和数量分布[18]。相关分析表明穗粒数与全生育期、株高和穗长呈极显著正相关,与有效穗率呈极显著负相关,相关系数的绝对值均在0.40~0.69,达到中度相关。本研究5个类群粳稻资源的表型性状有明显差异,且每个类群中粳稻资源的数量不同,第Ⅱ类只有中楼1号、BG1、兴图3份种质,具有大穗、大粒和高结实率等特点;第Ⅴ类群只有中大菲、云冷22号2份种质,具有生育期偏长、植株高大、穗大、粒多、小粒等特点。这5份资源可作为高产育种亲本材料。本研究结果可为粳稻育种和基础研究提供新种质。
3.2 粳稻资源综合性状评价
水稻种质资源是水稻新品种培育和重要农艺性状遗传研究的基础材料,优特异水稻种质资源的遗传改良和创新利用是提高水稻生产能力的直接有效的措施。福建省历史上稻作主要以种植籼稻为主,亦有粳稻和糯稻。据调查20世纪50年代福建省征集到约
3948 份稻种资源中,籼稻占79.6%,粳稻占20.4%[19],在福建省生产上大面积利用的粳稻品种基本没有。而近年来随着运输成本的增加,“东北大米”的输出正成为发展的“瓶颈”,福建省发展粳稻生产意义重大。根据各主成分的累积贡献率得到前4个主成分的权重,计算得到140份稻种资源的综合评价D值,筛选出排名前10位的资源分别是当育5号、奥村学、中大菲、中花15号、云冷22号、BG1、农虎禾、毕梗42、兴图、G珍汕9713 。新引进的140份粳稻资源不仅丰富福建稻种资源基因库,且经综合评价这10份资源在综合农艺性状及产量上具有一定优势,对福建粳稻育种亲本的选配、品种改良等应用研究具有重要意义。 -
![]()
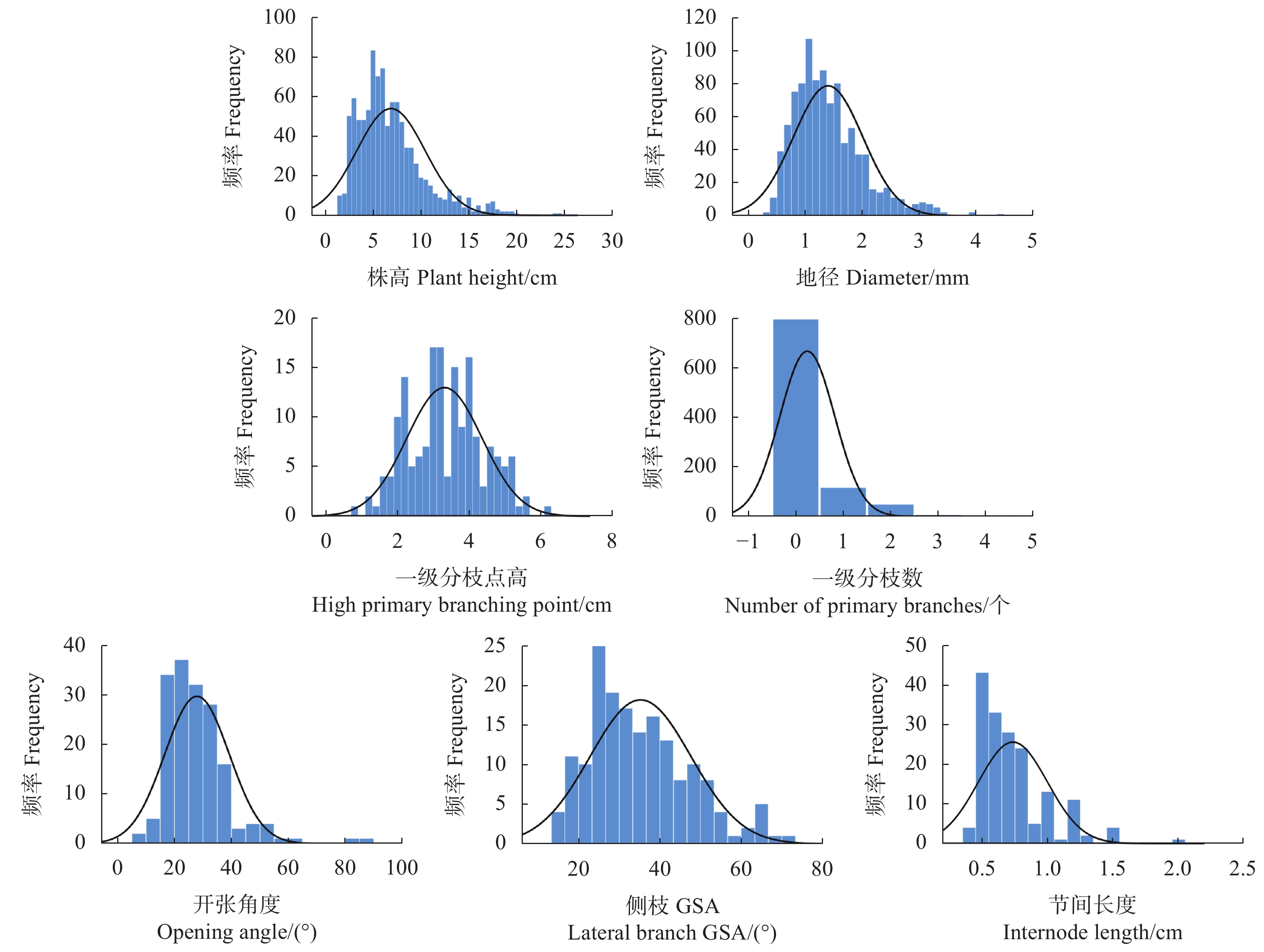
图 2 QC×FJ群体杂交后代株型性状的频率分布
Figure 2. Frequency distribution of phenotype traits in QC×FJ population
![]()
图 3 FJ×QC群体杂交后代株型性状的频率分布
Figure 3. Frequency distribution of phenotype traits in FJ×QC population
表 1 福建紫薇与‘千层绯雪’的杂交结实率
Table 1 Seed setting rates of L. limii ×'Qianceng Feixue' hybrids
母本×父本
Maternal×
Paternal杂交数量
Flower number
of crosses结实数
Fruit number结实率
Fruiting
rate/%出苗数
Seedling
number‘千层绯雪’×福建紫薇
‘Qianceng Feixue’×L.limii371 280 75.5 963 福建紫薇ב千层绯雪’
L.limiiבQianceng Feixue’17 17 100.0 66 合计 Total 388 297 76.5 1029  下载: 导出CSV
下载: 导出CSV
表 2 QC×FJ群体的描述性统计
Table 2 Descriptive statistics of QC×FJ population
性状
Traits最大值
Maximum最小值
Minimum均值
Average标准差
SD偏度
Skewness峰度
Kurtosis变异系数
CV/%株高
Plant height/cm24.50 1.50 6.87 3.57 1.32 2.16 51.97 地径
Diameter/mm4.44 0.30 1.40 0.61 1.09 1.55 43.57 一级分枝点高度
Height at first branching point/cm6.20 0.80 3.31 1.05 0.11 0.19 31.72 一级分枝数
Number of first-order branches4.00 0.00 0.24 0.58 2.59 6.84 240.42 开张角度
Opening angle/(°)87.56 8.20 27.90 11.35 1.89 6.43 40.68 侧枝GSA
Lateral branch GSA/(°)72.25 15.35 35.26 12.42 0.71 0.05 35.22 节间长度
Internode length/cm2.0 0.40 0.73 0.26 1.58 3.23 35.62
下载: 导出CSV
表 3 FJ×QC群体的描述性统计
Table 3 Descriptive statistics of FJ×QC population
性状
Traits最大值
Maximum最小值
Minimum均值
Average标准差
SD偏度
Skewness峰度
Kurtosis变异系数
CV/%株高
Plant height/cm22.0 2.0 7.54 4.08 1.06 2.19 54.11 地径
Diameter/mm2.98 0.44 1.40 0.60 0.91 0.15 42.86 一级分枝点高度
Height at first branching point/cm5.8 0.5 2.41 1.39 0.99 0.97 57.68 一级分枝数
Number of first-order branches/each2 0 0.26 0.51 1.84 2.69 196.15 开张角度
Opening angle/(°)59.69 3.03 27.34 19.70 −1.38 1.121 72.06 侧枝GSA
Lateral branch GSA/°48.35 10.9 24.65 11.33 0.83 −0.10 45.96 节间长度
Internode length/cm54.8 17.3 33.40 13.39 0.38 −1.05 40.08
下载: 导出CSV
表 4 株型性状的相关性分析(QC×FJ)
Table 4 Correlation between phenotype traits (QC×FJ)
株高
Plant height地径
Diameter一级分枝点高度
Height at first branching point一级分枝数
Number of first-order branches开张角度
Opening angle侧枝GSA
Lateral branch GSA地径
Diameter0.821** 一级分枝点高度
Height at first branching point0.317** 0.254** 一级分枝数
Number of first-order branches0.100 0.169* −0.019 开张角度
Opening angle0.456** 0.441** 0.207** 0.400** 侧枝GSA
Lateral branch GSA0.524** 0.480** 0.244** 0.027 0.578** 节间长度
Internode length0.743** 0.701** 0.195* 0.036 0.377** 0.534** *表示显著相关(P<0.05),**表示极显著相关(P<0.01)。表5同。
* indicates significant correlation (P<0.05), ** indicates extremely significant correlation (P<0.01). Same for Table 5.
下载: 导出CSV
表 5 株型性状的相关性分析(FJ×QC)
Table 5 Correlation between phenotypic traits (FJ×QC)
株高
Plant height地径
Diameter一级分枝点高度
Height at first branching point一级分枝数
Number of first-order branches开张角度
Opening angle侧枝GSA
Lateral branch GSA地径
Diameter0.791** 一级分枝点高度
Height at first branching point−0.169 −0.107 一级分枝数
Number of first-order branches−0.141 0.076 0.725** 开张角度
Opening angle0.192 0.367 0.398 0.585* 侧枝GSA
Lateral branch GSA0.413 0.475 0.177 0.170 0.718** 节间长度
Internode length0.639* 0.819** −0.118 −0.038 0.476 0.427
下载: 导出CSV
表 6 不同遗传模型下QC×FJ 群体的 AIC 值
Table 6 AIC of QC×FJ population by different genetic models
模型
Model株高
Plant height地径
Diameter开张角度
Opening angle侧枝GSA
Lateral branch GSA一级分枝点高度
Height at first branching point0MG 5188.884 391.072 1303.792 1334.032 506.3505 1MG-AD 4899.472 300.2864 1250.446 1313.571 503.0956 1MG-A 4942.795 306.6786 1248.605 1312.763 501.3233 1MG-EAD 5032.944 335.7514 1282.295 1318.325 506.3529 1MG-NCD 4993.272 331.1677 1287.728 1321.534 509.4304 2MG-ADI 4950.868 325.7444 1278.015 1314.864 515.8107 2MG-AD 4871.614 294.8911 1245.498 1305.003 497.1486 2MG-A 4910.321 302.0399 1249.31 1314.201 510.264 2MG-EA 4888.363 295.9231 1246.738 1303.926 491.0738 2MG-CD 5192.889 395.0742 1307.791 1338.033 510.3601 2MG-EAD 5190.889 393.0743 1305.791 1336.033 508.3601 加粗值代表最小 AIC 值;MG 代表主基因模型;A 代表加性效应;D 代表显性效应; I 代表上位性 效应;E 代表相等;N 代表负向;C 代表完全。表7同。
Data in bold: minimum AIC; MG: major gene model; A: additive effect; D: dominant effect; I: epistatic interaction; E: equal; N: negative; C: complete. Same for Table 7.
下载: 导出CSV
表 7 不同遗传模型下FJ×QC群体的 AIC 值
Table 7 AIC of FJ×QC population under different genetic models
模型
Model地径
Diameter开张角度
Opening angle0MG 123.9251 118.3907 1MG-AD 109.1332 118.0483 1MG-A 114.9762 118.7385 1MG-EAD 115.9589 119.0364 1MG-NCD 110.7568 119.0415 2MG-ADI 110.3914 122.8118 2MG-AD 100.7582 106.6611 2MG-A 111.5877 118.2622 2MG-EA 107.1847 112.1628 2MG-CD 127.9204 122.358 2MG-EAD 125.9204 120.358
下载: 导出CSV
表 8 QC×FJ、FJ×QC 群体入选模型的适合性检验
Table 8 Fitting of QC×FJ and FJ×QC population inclusion models
群体
Group性状
Traits模型
ModelU12 U22 U32 nW2 Dn QC×FJ 株高
Plant height2MG-AD 0.0251 (0.8741 )0.0453 (0.8314 )0.0565 (0.8121 )0.0393 (0.9369 )0.0206 (0.7996 )地径
Diameter2MG-AD 0.0004 (0.9851 )0.0005 (0.9822 )0.0003 (0.9866 )0.012(1) 0.0107 (0.9998 )开张角度
Opening angle2MG-AD 0.0015 (0.9692 )0.0004 (0.9927 )0.0127 (0.9103 )0.0077 (1.0008 )0.0184 (1)侧枝GSA
Lateral branch GSA2MG-EA 0.0008 (0.9776 )0.0013 (0.971)0.0647 (0.7992 )0.0206 (0.9964 )0.032(0.993) 一级分枝点高度
Height at first branching point2MG-EA 0.0014 (0.9704 )0.001( 0.9748 )0.0003 (0.986)0.064( 0.7929 )0.0621 (0.5055 )FJ×QC 地径
Diameter2MG-AD 0.0013 (0.9708 )0.0017 (0.9674 )0.0005 (0.9828 )0.0063 (1.0046 )0.0267 (1)开张角度
Opening angle2MG-AD 0.0217 (0.883)0.0142 (0.9051 )0.0087 (0.9257 )0.0152 (0.9996 )0.1078 (0.9871 )U12、 U22 和 U32 为均匀性检验,nW2 为 Smirnov 检验,Dn 为 Kolmogorov 检验,括号内为概率值。
U12, U22 and U32: Uniformity tests; nW2: Smirnov test; Dn: Kolmogorov test; data in brackets are probability.
下载: 导出CSV
表 9 QC×FJ、FJ×QC(一年生)群体各性状在最适模型下的遗传参数
Table 9 Genetic parameters of traits in QC×FJ and FJ×QC population under best fit models
群体
Group性状
Traitsda db ha hb i jab jba l h2mg /% QC×FJ 株高
Plant height3.5662 1.5173 −1.452 −0.782 0 地径
Diameter0.3965 0.1399 −0.226 −0.29 0 开张角度
Opening angle8.4046 6.8204 −3.912 −3.831 0 侧枝GSA
Lateral branch GSA8.8543 23.802 一级分枝点高度
Height at first branching point0.6373 63.933 FJ×QC 地径
Diameter0.6005 0.3401 −0.238 −0.257 91.1643 开张角度
Opening angle10.415 7.135 −1.855 −3.439 95.6964 da:第 1 对主基因加性效应;db:第 2 对主基因加性效应;ha:第 1 对主基因显性效应;hb:第 2 对主基因显性效应;i:加×加效应;jab:加×显应;jba:显×加效应;l:显×显效应;h2mg:主基因遗传率。
da: 1st major-gene additive effect; db: 2nd major-gene additive effect; ha: 1st major-gene dominant effect; hb: 2nd major-gene dominant effect; i: additivity × additivity effect; jab: additivity × dominance effect; jba: dominance × additivity effect; l: dominance × dominance effect; h2mg: major gene heritability.
下载: 导出CSV
-
[1] 中国科学院中国植物志编辑委员会. 中国植物志-第七十三卷,第二分册[M]. 北京:科学出版社,1983. [2] EGOLF D R,ANDRICK A O. The Lagerstroemia handbook/checklist:A guide to crapemyrtle cultivars[J]. USA:American Association of Botanical Gardens and Arboreta,1978.
[3] 王金凤,柳新红,陈卓梅. 紫薇属植物育种研究进展[J]. 园艺学报,2013,40(9) :1795−1804. WANG J F,LIU X H,CHEN Z M. Research progress in breeding of Lagerstroemia plant[J]. Acta Horticulturae Sinica,2013,40(9) :1795−1804. (in Chinese)
[4] 王甜. 紫薇和川黔紫薇远缘杂交亲和性及种子萌发特性研究[D]. 长沙:中南林业科技大学,2022. WANG T. Studies on Compatibility and Seed Germination Characteristics of the Distant Hybridization of Lagerstroemia indica and Lagerstroemia excelsa (Dode) Chun[D]. Changsha:Central South University of Forestry & Technology,2022. (in Chinese)
[5] SEGURA V,CILAS C,COSTES E. Dissecting apple tree architecture into genetic,ontogenetic and environmental effects:Mixed linear modelling of repeated spatial and temporal measures[J]. New Phytologist,2008,178(2) :302−314.
[6] SOCQUET-JUGLARD D,CHRISTEN D,DEVÈNES G,et al. Mapping architectural,phenological,and fruit quality QTLs in apricot[J]. Plant Molecular Biology Reporter,2013,31(2) :387−397.
[7] LIU R Z,AI N J,ZHU X X,et al. Genetic analysis of plant height using two immortalized populations of “CRI12 × J8891” in Gossypium hirsutum L[J]. Euphytica,2014,196(1) :51−61. DOI: 10.1007/s10681-013-1013-0
[8] ZHANG Y G,ZHU J,DAI H Y. Characterization of transcriptional differences between columnar and standard apple trees using RNA-seq[J]. Plant Molecular Biology Reporter,2012,30(4) :957−965.
[9] PETERSEN R,KROST C. Tracing a key player in the regulation of plant architecture:The columnar growth habit of apple trees (Malus × domestica) [J]. Planta,2013,238(1) :1−22. DOI: 10.1007/s00425-013-1898-9
[10] 陈卓梅,沈鸿明,张冬林,等. 从美国专利品种看中国紫薇育种发展趋势[J]. 浙江林业科技,2018,38(1) :49−61. CHEN Z M,SHEN H M,ZHANG D L,et al. Patented cultivars of Lagerstroemia in the US and trends of breeding in China[J]. Journal of Zhejiang Forestry Science and Technology,2018,38(1) :49−61. (in Chinese)
[11] 国家林业局科技发展中心(国家林业局植物新品种保护办公室) . 中国林业植物授权新品种-2016[M]. 北京:中国林业出版社,2017. [12] 胡琦,何金儒,潘会堂. 紫薇新品种[J]. 中国花卉园艺,2022(6) :32−33. DOI: 10.3969/j.issn.1009-8496.2022.6.zghhyy202206009 HU Q,HE J R,PAN H T. New Cultivar of Lagerstroemia indica[J]. China Flowers & Horticulture,2022(6) :32−33. (in Chinese) DOI: 10.3969/j.issn.1009-8496.2022.6.zghhyy202206009
[13] 赵静媛,陈发棣,滕年军,等. 地被菊匍匐性的遗传分析与RAPD标记研究[J]. 中国农业科学,2009,42(2) :734−741. ZHAO J Y,CHEN F D,TENG N J,et al. Genetic analysis and RAPD marker of creeping habits in ground-cover Chrysanthemum[J]. Scientia Agricultura Sinica,2009,42(2) :734−741. (in Chinese)
[14] 刘阳. 紫薇微卫星标记开发及矮化性状的分子标记[D]. 北京:北京林业大学,2013 LIU Y. Development of Microsatellite of Lagerstroemia and Molecular Markers of Dwarf Trait[D]. Beijing:Beijing Forestry University,2013. (in Chinese)
[15] 石俊,陈之琳,秦波,等. 紫薇匍匐平展性状相关表型遗传分析[C]/ /张启翔. 中国观赏园艺研究进展2016. 北京:中国林业出版社,2016. [16] 焦垚. 大花紫薇与紫薇杂交后代重要观赏性状遗传分析[D]. 北京:北京林业大学,2017 JIAO Y. Phenotypic and Genetic Analysis of Ornamental Traits in Hybrids of Lagerstroemia indica and L. speciosa[D]. Beijing:Beijing Forestry University,2017. (in Chinese)
[17] 黄冬贤,黎可华,黎明星,等. 紫薇的整形修剪及其园林应用[J]. 广东园林,2010,32(4) :65−67. DOI: 10.3969/j.issn.1671-2641.2010.04.016 HUANG D X,LI K H,LI M X,et al. Pruning technology and landscape application of Lagerstroemia indica[J]. Guangdong Landscape Architecture,2010,32(4) :65−67. (in Chinese) DOI: 10.3969/j.issn.1671-2641.2010.04.016
[18] YE Y J,WU J Y,FENG L,et al. Heritability and gene effects for plant architecture traits of crape myrtle using major gene plus polygene inheritance analysis[J]. Scientia Horticulturae,2017,225:335−342.
[19] 姜珊,易星湾,徐庭亮,等. 月季花瓣数量遗传分析[J]. 植物科学学报,2021,39(2) :142−151. DOI: 10.11913/PSJ.2095-0837.2021.20142 JIANG S,YI X W,XU T L,et al. Genetic analysis of petal number in Rosa[J]. Plant Science Journal,2021,39(2) :142−151. (in Chinese) DOI: 10.11913/PSJ.2095-0837.2021.20142
[20] 吴元奇,唐兰,邱贵兰,等. 一份矮生玉米株高主基因+多基因遗传模式分析[J]. 四川农业大学学报,2022,40(3) :353−361. WU Y Q,TANG L,QIU G L,et al. Genetic pattern analysis of the height main gene and multigene of A dwarf maize[J]. Journal of Sichuan Agricultural University,2022,40(3) :353−361. (in Chinese)
[21] 王靖天,张亚雯,杜应雯,等. 数量性状主基因+多基因混合遗传分析R软件包SEA v2.0[J]. 作物学报,2022,48(6) :1416−1424. WANG J T,ZHANG Y W,DU Y W,et al. SEA v2.0:An R software package for mixed major genes plus polygenes inheritance analysis of quantitative traits[J]. Acta Agronomica Sinica,2022,48(6) :1416−1424. (in Chinese)
[22] 张向前,刘景辉,齐冰洁,等. 燕麦种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报,2010,11(2) :168−174. ZHANG X Q,LIU J H,QI B J,et al. Cluster diversity analysis of the main agronomic traits in oat germplasm[J]. Journal of Plant Genetic Resources,2010,11(2) :168−174. (in Chinese)
[23] 秦波,李素珍,胡玲,等. 基于转录组的紫薇SSR分子标记开发[C]//中国观赏园艺研究进展2018. 哈尔滨,2018:526–532. [24] 郭长花,康向阳. 树木发育中的阶段转变研究进展[J]. 生物技术通讯,2008,19(5) :784−786. GUO C H,KANG X Y. Progress in phase change during tree development[J]. Letters in Biotechnology,2008,19(5) :784−786. (in Chinese)
[25] GREENWOOD M S. Phase change in loblolly pine:Shoot development as a function of age[J]. Physiologia Plantarum,1984,61(3) :518−522. DOI: 10.1111/j.1399-3054.1984.tb06366.x
[26] 戴思兰. 园林植物育种学[M]. 北京:中国林业出版社,2007. [27] 叶福民,马策,张晓波,等. 多头切花秋菊杂交F1群体观赏性状主基因+多基因混合遗传分析[J]. 辽宁农业科学,2022(6) :7−12. YE F M,MA C,ZHANG X B,et al. Heterosis of ornamental characters and mixed inheritance of major gene and polygene in hybrid progenies of cut Chrysanthemum[J]. Liaoning Agricultural Sciences,2022(6) :7−12. (in Chinese)
[28] 张琳,郭丽丽,郭大龙,等. 牡丹杂交F1代性状分离规律及混合遗传分析[J]. 南京林业大学学报(自然科学版) ,2018,42(6) :51−60. ZHANG L,GUO L L,GUO D L,et al. Separation analysis and mixed genetic analysis of phenotypic traits in F1 progenies of tree peony[J]. Journal of Nanjing Forestry University (Natural Sciences Edition) ,2018,42(6) :51−60. (in Chinese)




计量
- 文章访问数: 66
- HTML全文浏览量: 10
- PDF下载量: 12
