

Infestation, Population Dynamics, and Host Preference of Thrips on Flowering Pummelo Plants
-
摘要:目的
明确福建蜜柚花期蓟马的为害特征、种类组成、种群时空动态及寄主选择性,为针对性制定防控措施提供科学依据。
方法选取漳州市平和县传统蜜柚果园为调查地点,以红肉蜜柚、白肉蜜柚和葡萄柚等蜜柚果树为对象,采集蜜柚花上的蓟马及被为害幼果,利用形态学特征对蓟马进行鉴定,记录不同采样时间、海拔高度、蜜柚品种的蓟马种类及数量,检测幼果营养品质。
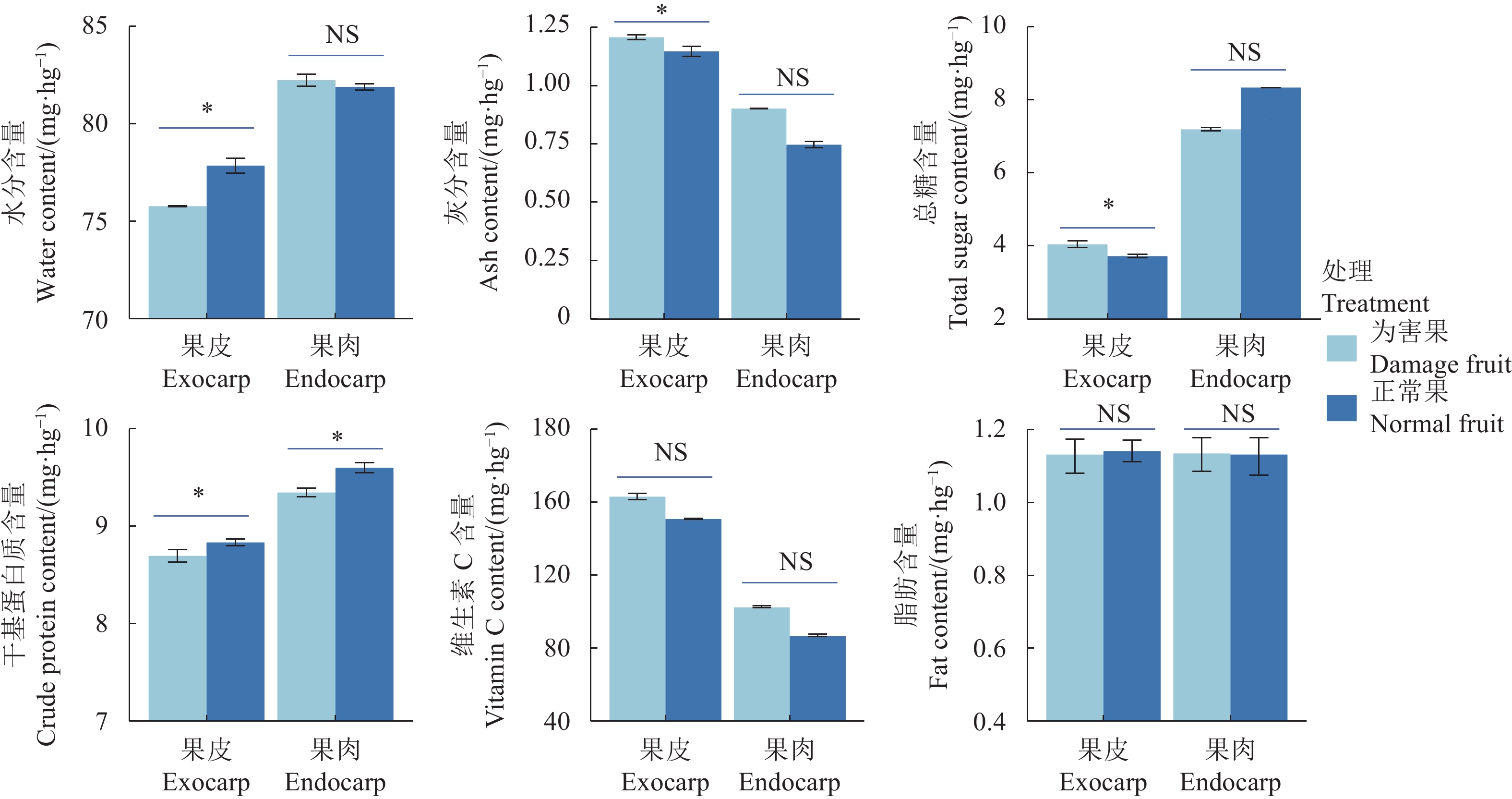
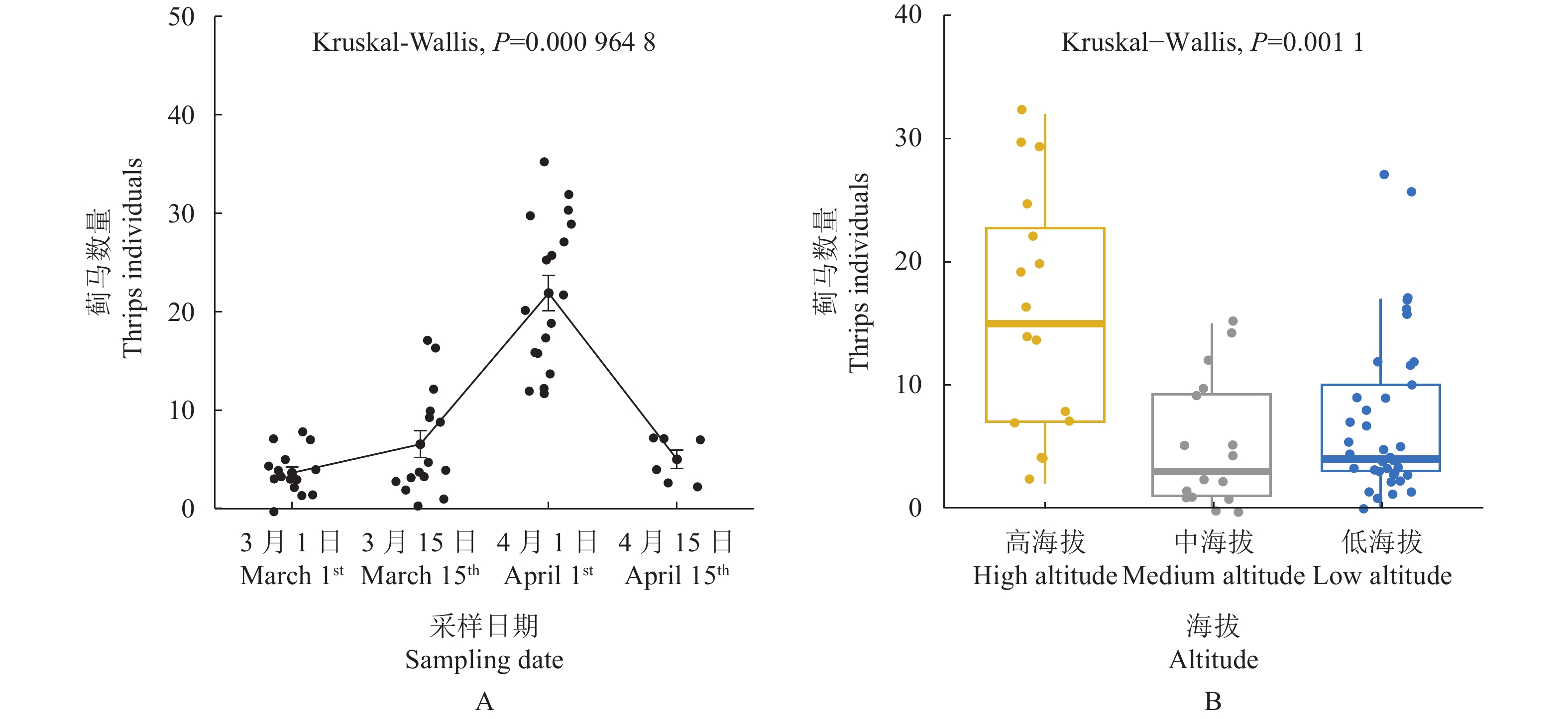
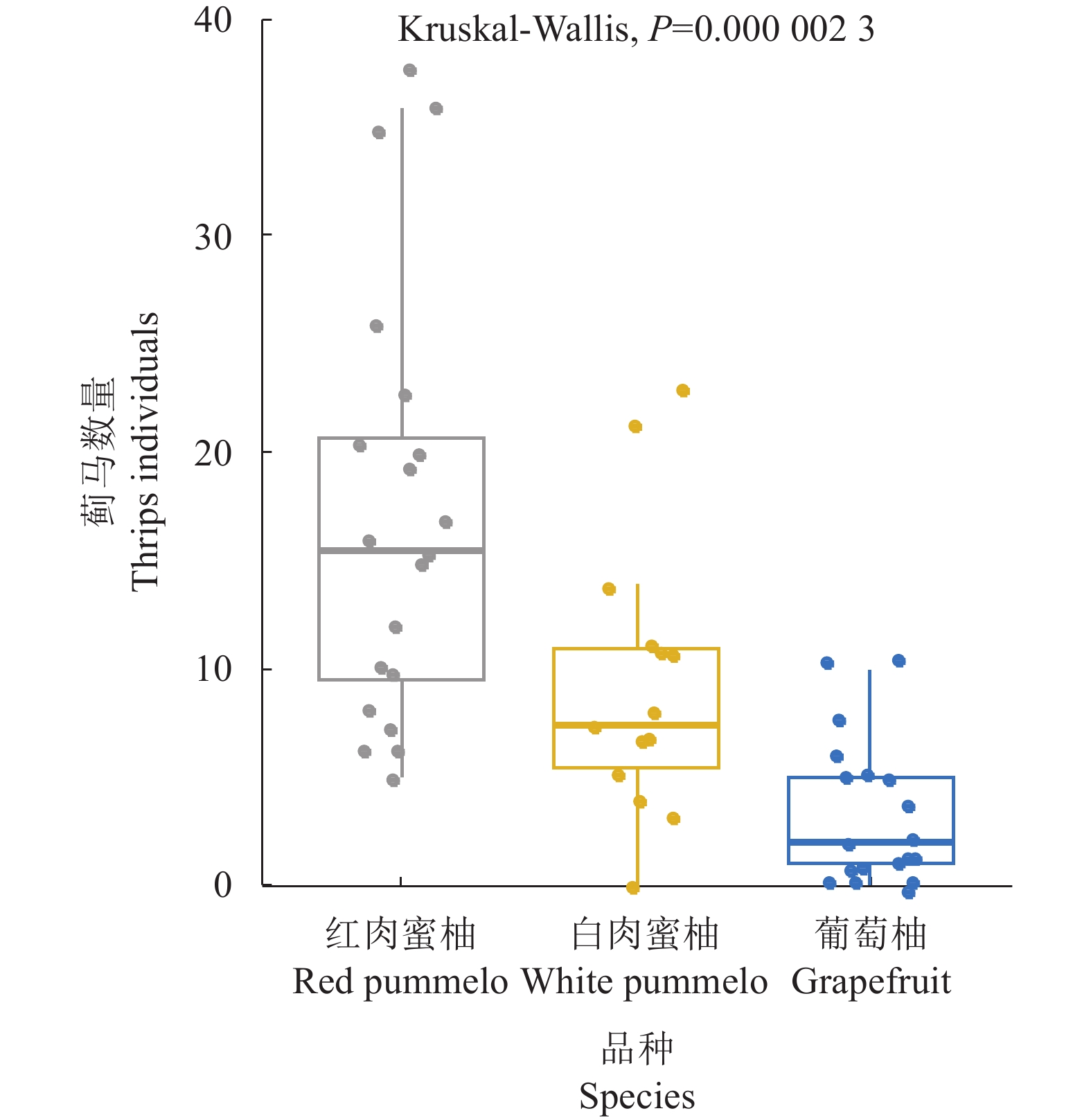
结果蜜柚花期蓟马为害导致果蒂周围形成环状疤痕,而且影响幼果果皮及果肉的营养成分,果皮的水分和干基蛋白质含量显著降低,灰分和总糖含量显著升高,而果肉仅有干基蛋白质显著降低。蜜柚花期蓟马属混合种群,由黄胸蓟马(Thrips hawaiiensis)、花蓟马(Frankliniella intonsa)、茶黄蓟马(Scirtothrips dorsali)、杜鹃蓟马(Thrips anderwsi)等组成,其优势种为黄胸蓟马,占比为70.8%~92.0%。蜜柚花期蓟马种群数量随时间及海拔高度而改变,3月下旬至4月上旬种群数量达高峰区,为20头·株−1以上,且高海拔种植园蜜柚蓟马发生数量显著高于中低海拔种植园。蜜柚花期蓟马对不同品种蜜柚的嗜好选择存在显著差异,传统品种红肉蜜柚、白肉蜜柚上蓟马发生数量显著高于葡萄柚等新引进品种。
结论蜜柚花期蓟马不仅影响柚子的外观品质,也影响其内在营养品质。花期蓟马群落中优势种为黄胸蓟马,需重点防控,在针对性防控措施制定时应考虑开花时期、海拔高度及蜜柚品种对蓟马种群数量的影响。
Abstract:ObjectiveInfestation, population dynamics, and host preferences of thrips on pummelo plants during flowering period were studied for an effective pest control.
MethodTraditionally managed pummelo orchards in Pinghe County, Zhangzhou City, Fujian that grew varieties including the conventional red and white pummelos and the newly introduced grapefruit were surveyed under this study. On sampling dates during pummelo flowering season, varieties of the infested pummelo plants and altitude of the plantation site as well as population of the invading thrips were recorded. Young pummelo fruits damaged by the pests were collected for nutritional quality determination.
ResultsThrip infestation on pummelos was generally manifested with circular scars around the fruit stem and reduced nutrients in the peel and pulp of the affected young fruits. The moisture and protein contents in the peels were significantly decreased, but the ash and total sugar contents increased, while the protein in the pulp slightly declined. In the pummelo flowering stage, the thrip population was mainly comprised of Thrips hawaiensis, Frankliniella intonsa, Scirtothrips dorsalis, andT. anderwsi with T. hawaiensis being the dominant species that contributed 70.8%–92.0% of total. The population peaked in early April and significantly affected by both plant variety and plantation elevation. For instance, the pummelo orchards located in high altitude encountered significantly greater numbers of thrips, and the traditional red and white pummelos tended to attract significantly more thrips than the newly introduced varieties, such as grapefruits.
ConclusionThe flowering pummelo plants infested by thrips produced unappealing fruits with inferior nutritional quality. Being the dominant infestation species,T. hawaiensis should be prioritized in the prevention and control programs at the orchards. It would also be prudent to include pesticide application time, altitude of plantation location, and species of pummelo involved into consideration for the implementation.
-
Keywords:
- Pummelo /
- thrips /
- damage by infestation /
- population dynamics /
- host preference
-
0. 引言
【研究意义】大豆胞囊线虫(H. glycines Ichinohe,Soybean cyst nematode,SCN)是威胁全球大豆正常生长的重要病原物,每年在全球范围内造成数十亿美元的经济损失,防控任务艰巨。深入挖掘抗病基因,解析抗病基因的作用机制对培育抗线虫的大豆品种具有重要意义。【前人研究进展】木质素(lignin)是存在于维管植物细胞壁内的一种重要的次生代谢物,在植物体内的含量丰富,仅次于纤维素[1-2]。它可以提高植物抵抗外界不良环境的能力[3],并且在水分运输、抵抗病原物入侵以及损伤修复等方面具有极其重要的作用[4]。方东鹏等[5]研究发现,木质素含量的增加能够提高野生大豆对疫霉菌(Phytophthora sojae)的抗性。颜清上等[6]研究结果表明大豆接种大豆胞囊线虫(H. glycines)4号生理小种后,抗性品种灰皮支黑豆和元钵黑豆中的类黄酮和木质素含量增加,而在感病品种鲁豆1号中含量下降,据此推测出根部的木质化作用是抗病品种对大豆胞囊线虫抗性的一种机制。咖啡酰辅酶A-O-甲基转移酶(CCoAOMT)是木质素生物合成过程中重要的S-腺苷-L-甲硫氨酸(SAM)甲基转移酶[7],生物碱和黄酮等化合物能够被其催化[8]。CCoAOMT相对分子量小,反应时依靠Mg2+进行激活,再以咖啡酰辅酶A作为底物,催化反应物SAM上的甲基转移到木质素上,形成阿魏酰辅酶A[9-10]。王宇光等[11]研究认为甜菜CCoAOMT基因可以提高植株的抗旱及耐盐特性,转CCoAOMT基因的甜菜植株的木质素含量显著增加。在盐诱导条件下,水稻中CCoAOMT基因表达上调[12],干旱胁迫可激活玉米中CCoAOMT的活性,推测其抗旱性与木质素合成有密切关系[13]。1989年,Kühnl等第一次在胡萝卜中发现了CCoAOMT [14],此后Pakusch又在欧芹中发现了CCoAOMT[15]。随着研究的深入,人们发现CCoAOMT存在多基因家族现象[16],并陆续在拟南芥[17]、绿竹[3]、棉花[18]、甜菜[19]、茶树[20]等植物中分离得到CCoAOMT。【本研究切入点】有关大豆CCoAOMT基因的研究甚少。在前期的研究中,我们利用蛋白质组学技术筛选出与大豆胞囊线虫3号生理小种抗性密切相关的差异蛋白咖啡酰辅酶A-O-甲基转移酶[21]。据此推测木质素代谢途径与大豆抗胞囊线虫存在相关性,根部的木质化作用是其对胞囊线虫抗性的一种机制。然而有关灰皮支黑豆CCoAOMT 基因的相关研究还有待深入进行。【拟解决的关键问题】以抗线大豆灰皮支黑豆为试验材料,克隆其CCoAOMT基因,并对该基因及其编码的氨基酸序列进行生物信息学分析,以期为进一步研究CCoAOMT基因在大豆抗胞囊线虫中的功能提供理论支撑。
1. 材料与方法
1.1 供试大豆
灰皮支黑豆(ZDD 2315),高抗大豆胞囊线虫病品种。
1.2 试验方法
1.2.1 大豆根系总RNA的提取和质量检测
将灰皮支黑豆的幼嫩根系用自来水冲洗干净,使用AXYGEN公司的Total RNA Miniprep Kit提取根系的总RNA,然后利用1%琼脂糖凝胶电泳检测RNA的质量,最后再利用Nanodrop紫外分光光度计测定RNA的浓度。
1.2.2 大豆CCoAOMT基因的PCR扩增
根据TIANGEN公司的cDNA合成系统,以Oligo(dT)15为引物,利用Quant Reverse Transcriptase将提取的总RNA合成高质量的cDNA。逆转录反应体系:RNA模板 2 μL,10×RT缓冲液 2 μL,dNTP混合液(2.5 mmol·L−1)4 μL,Oligo(dT)15(10 μmol·L−1)2 μL,RNase抑制剂(10 U·μL−1)1 μL,Quant Reverse Transcriptase 1 μL,无RNase的水8 μL。
RT-PCR反应体系:cDNA模板2 μL,10 μmol·L−1 CCA-RTS 1 μL,10 μmol·L−1 CCA-RTA 1 μL,2×Taq PCR MasterMix 12.5 μL,灭菌ddH2O 8.5 μL。RT-PCR反应程序:预变性94 ℃ 2 min;变性94 ℃ 30 s,退火58 ℃ 1 min,72 ℃延伸 2 min,30个循环;72 ℃ 延伸10 min。DNA marker 购自TIANGEN公司,PCR引物由TaKaRa公司合成,序列:CCA-RTS: 5′-CTAGTCCCAAAGAGCTAACAA-3′;CCA-RTA: 5′-CCACCATTCACGCAACCT-3′。
1.2.3 RT-PCR产物的克隆及测序
利用AXYGEN公司的DNA Gel Extraction Kit回收并纯化PCR产物,利用克隆载体pGEM-T Easy将其克隆并转化到大肠杆菌DH5α,连接体系:回收DNA片段 3 μL,T-easy载体 1 μL,T4 DNA Ligase 1 μL,2×Rapid Ligation Buffer 5 μL。挑取阳性克隆、提取质粒,送至大连TaKaRa公司测序。
1.2.4 大豆CCoAOMT基因的生物信息学分析
(1)同源性分析 利用TBtools对克隆获得的大豆CCoAOMT基因的开放阅读框(Open Reading Frame, ORF)进行预测,利用DANMAN6.0将其翻译成氨基酸序列[22],在NCBI网站上采用BLASTp程序对氨基酸序列进行保守结构域预测及同源性分析。
(2)理化性质分析 利用在线程序ProtParam(https://web.expasy.org/protparam/)进行大豆及其他物种的CCoAOMT蛋白的氨基酸组成和理化性质分析[23];利用在线程序ProtScale(https://web.expasy.org/protscale/)进行亲、疏水性分析[24]。
(3)亚细胞定位、拓扑结构、跨膜区和信号肽预测 利用在线程序CELLO(http://cello.life.nctu.edu.tw/)对GmCCoAOMT蛋白进行亚细胞定位预测[25];利用PROTTER(http://wlab.ethz.ch/protter/#)对GmCCoAOMT蛋白的拓扑结构进行分析,判断该蛋白是胞内蛋白还是分泌蛋白;利用TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)对GmCCoAOMT跨膜区域进行预测;利用SignalP-5.0(http://www.cbs.dtu.dk/services/SignalP/)进行信号肽预测。
(4)蛋白结构预测 利用在线工具prabi(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)对大豆CCoAOMT蛋白进行二级结构预测;利用Swiss Model(https://swissmodel.expasy.org/interactive)对大豆CCoAOMT蛋白进行三级结构预测。
(5)系统进化分析 利用MEGA-7.0中的MUSCLE进行多重序列比对,并利用最大似然法(Maximum Likelihood,ML)构建进化树[26]。氨基酸替换模型为JTT+G,利用Bootstrap重复1000次评估得到各节点的支持率,Partial deletion值设置为95%。
2. 结果与分析
2.1 大豆根系总RNA的提取和质量检测
灰皮支黑豆根系的总RNA经1%琼脂糖凝胶电泳检测,结果显示18S、28SrRNA完整,表明RNA的完整性良好,未发生降解(图1)。紫外分光光度计法检测RNA的OD260/OD280的3次平均值为1.98,表明RNA的纯度和质量均符合试验要求。
2.2 大豆CCoAOMT基因的克隆与序列分析
以灰皮支黑豆根系的cDNA为模板,扩增到1条长度为848 bp的CCoAOMT基因片段(图2),将其命名为GmCCoAOMT,其所编码的蛋白质为GmCCoAOMT。该基因包含1个长度为741 bp的开放阅读框(图3)。对灰皮支黑豆CCoAOMT基因编码氨基酸的理化性质进行分析表明:GmCCoAOMT基因编码1个由246个氨基酸组成的蛋白,该蛋白由3926个原子构成,分子式为C1244H1985N325O362S10,分子量为27.6 kDa,氨基酸组分中含量最多的是亮氨酸(Leu),占14.6%。该蛋白的负电荷残基(Asp + Glu)数为34个,正电荷残基(Arg + Lys)为28个,理论等电点为5.67;该蛋白的不稳定指数为40.55,据此可将其归为不稳定蛋白;脂肪族指数为105.49,平均疏水性(GRAVY)为−0.169。
![]() 图 2 CCoAOMT基因的RT-PCR结果M: DNA marker;1、2: RT-PCR 产物Figure 2. RT-PCR on CCoAOMTM: DNA marker;1 and 2: RT-PCR products.
图 2 CCoAOMT基因的RT-PCR结果M: DNA marker;1、2: RT-PCR 产物Figure 2. RT-PCR on CCoAOMTM: DNA marker;1 and 2: RT-PCR products.利用NCBI的Conserved Domains对CCoAOMT的结构域进行预测,结果表明,GmCCoAOMT含有1个保守的AdoMet_MTases结构域和腺苷甲硫氨酸结合位点,属于AdoMet_MTases超家族(图4),E-value值为4.18e−153。亲疏水性预测显示(图5),在GmCCoAOMT蛋白中,亲水性氨基酸的数量多于疏水性氨基酸,由此可推测该蛋白属于亲水性蛋白;其中位于6位点的谷氨酸亲水性最强,位于96位点的苏氨酸疏水性最强。
![]() 图 3 大豆CCoAOMT基因全长序列及其编码的氨基酸序列Figure 3. Full-length cDNA and amino acid sequence of CCoAOMT in soybean
图 3 大豆CCoAOMT基因全长序列及其编码的氨基酸序列Figure 3. Full-length cDNA and amino acid sequence of CCoAOMT in soybean亚细胞定位预测结果表明,GmCCoAOMT蛋白位于细胞质(Cytoplasm)的可能性最大,分值为3.309。拓扑结构分析预测结果表明,GmCCoAOMT蛋白为胞内蛋白(图6)。TMHMM预测结果显示,GmCCoAOMT蛋白不含跨膜区域,表明该蛋白可能是一种非跨膜蛋白,SignalP信号肽预测结果显示,GmCCoAOMT基因编码的氨基酸不含信号肽。
2.3 大豆与其他豆科植物CCoAOMT蛋白的比较分析
2.3.1 氨基酸同源性分析
将本研究获得的大豆CCoAOMT编码的氨基酸序列与NCBI数据库中的其他豆科植物菜豆(Phaseolus vulgaris Linn.)、虎爪豆(Mucuna pruriens L.)、木豆[Cajanus cajan (Linn.) Millsp.]、密花豆(Spatholobus suberectus Dunn)、相思子(Abrus precatorius L.)、白羽扇豆(Lupinus albus)、豇豆[Vigna unguiculata (Linn.)Walp.]、赤豆[Vigna angularis (Willd.) Ohwi et Ohashi]、红车轴草(Trifolium pratense L.)以及鹰嘴豆(Cicer arietinum Linn.)的CCoAOMT蛋白氨基酸序列(登录号分别为AGV54319.1、RDX78299.1、XP_020208653.1、TKY55699.1、XP_027347195.1、KAE9597637.1、QCD81613.1、XP_017427550.1、PNY11893.1、NP_001351681.1)进行多重序列比对,结果如图7所示。可知大豆[Glycine max (Linn.) Merr.]与菜豆的CCoAOMT相似性最高,达89.02%,与赤豆、木豆、豇豆、密花豆、相思子、红车轴草以及鹰嘴豆的相似性分别为88.21%、88.62%、86.18%、84.55%、84.15%、79.01%和73.17%。
![]() 图 7 大豆与其他豆科植物CCoAOMT蛋白氨基酸多重序列比对Figure 7. Multiple alignment of amino acid sequences between CCoAOMTs in soybean and other legumes
图 7 大豆与其他豆科植物CCoAOMT蛋白氨基酸多重序列比对Figure 7. Multiple alignment of amino acid sequences between CCoAOMTs in soybean and other legumes2.3.2 蛋白理化性质及氨基酸组成分析
对上述11种豆科植物CCoAOMT蛋白的理化性质与氨基酸组成进行分析,结果(表1)表明:11种豆科植物CCoAOMT蛋白的相对分子量大小差异不明显,其中大豆CCoAOMT蛋白的相对分子量最低,为27.6 kDa,而其他豆科植物CCoAOMT蛋白的相对分子量均在27.8~28.0 kDa。11种豆科植物CCoAOMT蛋白的等电点在5.32~5.90,没有显著区别。11种豆科植物CCoAOMT蛋白的氨基酸数量为246~248个,其中菜豆、木豆、豇豆、密花豆、赤豆、相思子CCoAOMT蛋白的氨基酸数量与大豆一致,均为246个,白羽扇豆、鹰嘴豆、虎爪豆和红车轴草CCoAOMT蛋白的氨基酸数量比大豆稍多,分别为247、247、248和248个。从氨基酸的组成来看,除红车轴草外,其他10种豆科植物的CCoAOMT蛋白中酸性氨基酸与碱性氨基酸比例相差不多,均在14%~15%,脂肪族氨基酸所占比例均在80%以上,显著高于芳香族氨基酸。
表 1 大豆和其他豆科植物CCoAOMT蛋白的氨基酸组成和理化性质对比Table 1. Amino acid composition and physicochemical properties of CCoAOMTs in soybean and other plants序号
Serial number物种
Species登录号
Accession No.氨基酸数量
Number of
amino acids相对分子量
Relative molecular
mass/kDa理论等电点
Theoretical isoelectric
point比例
Percentage/%BA AC AR AL 1 大豆 Glycine max MW480860 246 27.6 5.67 14.2 13.8 7.3 84.6 2 菜豆 Phaseolus vulgaris AGV54319.1 246 27.8 5.32 13.8 14.6 8.5 83.4 3 虎爪豆 Mucuna pruriens RDX78299.1 248 28.0 5.51 14.1 14.2 7.6 84.8 4 木豆 Cajanus cajan XP_020208653.1 246 27.8 5.72 15.1 14.2 6.9 84.1 5 密花豆 Spatholobus suberectus TKY55699.1 246 27.9 5.47 13.8 14.2 8.1 84.6 6 相思子 Abrus precatorius XP_027347195.1 246 27.9 5.50 14.7 14.6 7.3 83.7 7 白羽扇豆 Lupinus albus KAE9597637.1 247 28.0 5.40 14.1 14.6 8.1 84.2 8 豇豆 Vigna unguiculata QCD81613.1 246 27.8 5.48 15.1 15.0 7.3 83.7 9 赤豆 Vigna angularis XP_017427550.1 246 27.8 5.90 14.2 13.4 8.5 83.0 10 红车轴草 Trifolium pratense PNY11893.1 248 27.9 5.43 12.4 12.9 7.2 86.0 11 鹰嘴豆 Cicer arietinum NP_001351681.1 247 28.0 5.69 15.0 14.6 8.1 84.6 BA:碱性氨基酸;AC:酸性氨基酸;AR:芳香族氨基酸;AL:脂肪族氨基酸。
BA: Basic amino acids; AC: acidic amino acids; AR: aromatic amino acids; AL: aliphatic amino acids.2.3.3 系统进化分析
利用MEGA-7.0软件对上述11种豆科植物CCoAOMT蛋白进行系统进化分析,利用最大似然法构建系统发生树,氨基酸替换模型为JTT+G。利用Bootstrap重复1000次评估得到各节点的支持率,Partial deletion值设置为95%。结果表明:大豆与豇豆、木豆、菜豆和赤豆CCoAOMT蛋白的进化关系较近,其次是相思子、密花豆、虎爪豆和红车轴草,而与白羽扇豆和鹰嘴豆的进化关系较远(图8)。
![]() 图 8 大豆与其他植物CCoAOMT蛋白的进化树Figure 8. Phylogenetic tree of CCoAOMT proteins in soybean and other plants
图 8 大豆与其他植物CCoAOMT蛋白的进化树Figure 8. Phylogenetic tree of CCoAOMT proteins in soybean and other plants2.4 大豆CCoAOMT蛋白二级结构和三级结构预测
利用prabi对大豆CCoAOMT蛋白的二级结构进行预测(图9),α-螺旋占44.72%,无规则卷曲占29.27%,延伸链占17.48%,β-折叠结构占7.72%;利用Swiss Model软件对大豆CCoAOMT蛋白的三级结构进行预测(图10),模型构建参照高粱CCoAOMT的晶体结构(SMTL ID : 5kva.1),GMQE值为0.84,Seq Identity为67.63%。拉式图验证表明,模型构建准确。
![]() 图 9 大豆CCoAOMT蛋白的二级结构预测Figure 9. Predicted secondary structure of soybean CCoAOMT protein
图 9 大豆CCoAOMT蛋白的二级结构预测Figure 9. Predicted secondary structure of soybean CCoAOMT protein![]() 图 10 大豆CCoAOMT蛋白的三维建模Figure 10. Predicted tertiary structure of soybean CCoAOMT protein
图 10 大豆CCoAOMT蛋白的三维建模Figure 10. Predicted tertiary structure of soybean CCoAOMT protein3. 讨论与结论
在病原物与寄主植物的相互作用过程中,病原物侵染寄主植物以后,植物细胞壁的木质化作用即木质素含量明显增加,这为阻止病原物对寄主植物的严重为害提供了有效的保护作用,是寄主植物抗性反应的一种特性[27]。木质素是植物体内一种重要的次生代谢物,在植物细胞中具有非常重要的生物学功能,它能够增强植物的韧性和机械强度,从而提高植物的抗逆性。参与木质素生物合成途径的有关酶类很多,咖啡酰辅酶A-O甲基转移酶是其中一种催化甲基化作用的重要的甲基转移酶[28-30],植物中木质素含量可以通过改变木质素生物合成途径中相关基因的表达来调节。陈夏鑫[31]从荸荠的球茎中克隆到全长为1302 bp的CCoAOMT基因,GenBank登录号为MG719240,该基因包含一个786 bp的最大开放阅读框,而大豆CCoAOMT基因包含一个741 bp的开放阅读框。陈凌娜[32]通过RT-PCR从巨龙竹中克隆到全长为777 bp的CCoAOMT基因,预测其蛋白结构含有丰富的α-螺旋和无规则卷曲,这与本研究预测的大豆CCoAOMT蛋白结构相似,均有大量的α-螺旋。刘鑫等[33]从砀山酥梨中克隆到长度为750 bp的CCoAOMT基因,该基因在拟南芥中过表达后可促进其次生壁发育及木质化,表明该基因在植物木质素合成中发挥重要的作用。李元玉[34]从菘蓝中克隆到CCoAOMT基因的cDNA序列,长度为1 098 bp,并对其表达特征进行分析,为培育抗病毒活性高的优良菘蓝品系奠定基础。有关于大豆CCoAOMT基因的结构特征及其在抗SCN中的功能还未见详细报道,因此,本研究克隆了大豆CCoAOMT基因的序列并对其进行分析。
本研究克隆获得的灰皮支黑豆CCoAOMT基因的cDNA序列全长为848 bp,GenBank登录号为MW480860,编码246个氨基酸,预测分子量为27.6 kDa,理论等电点为5.67。灰皮支黑豆CCoAOMT蛋白含有一个保守的AdoMet_MTases结构域,属于AdoMet_MTases超级家族,亚细胞定位预测结果显示该蛋白位于细胞质。马倩等[35]对紫花苜蓿的44个CCoAOMT家族基因进行预测,其中33个CCoAOMT蛋白定位于细胞质,11个定位于叶绿体。灰皮支黑豆与其他豆科植物的CCoAOMT蛋白序列相似度较高,与菜豆CCoAOMT蛋白序列相似性达89.02%;系统进化分析表明:大豆与豇豆、木豆、菜豆和赤豆CCoAOMT蛋白进化关系较近,而与白羽扇豆和鹰嘴豆进化关系较远。本研究利用生物信息学的方法,对大豆CCoAOMT基因进行了初步的分析和研究,下一步将对CCoAOMT基因的组织表达情况进行qRT-PCR分析,为进一步研究木质素代谢途径关键基因CCoAOMT在大豆抗胞囊线虫中的作用机理奠定基础,进而明确大豆CCoAOMT蛋白的功能。
-
![]()
图 1 蓟马在蜜柚花(A、B)和果实(C)上的为害状
Figure 1. Flower (A and B) and fruit (C) of pummelo plants infested by thrips
![]()
图 2 不同部位的蓟马为害果的营养品质变化
“*”表示差异显著(P <0.05),“NS”表示无显著差异(P >0.05)。
Figure 2. Change on nutrients in pummelo fruit parts due to thrip infestation
* indicates significant difference at P<0.05; NS indicates no significant difference at P>0.05.
![]()
图 4 蜜柚上优势蓟马种群数量在时间(A)和空间(B)上的动态
Figure 4. Temporal (A) and spatial (B) dynamics of dominant thrip population on pummelo plants
-
[1] 施清. 福建柑橘产业现状与发展方向 [J]. 东南园艺, 2014, 2(5):36−38. DOI: 10.3969/j.issn.1004-6089.2014.05.008 SHI Q. Status and development direction of citrus industry in Fujian Province [J]. Southeast Horticulture, 2014, 2(5): 36−38. (in Chinese) DOI: 10.3969/j.issn.1004-6089.2014.05.008
[2] 毛加梅, 岳建强, 高俊燕. 我国果树蓟马研究进展 [J]. 江西农业学报, 2013, 25(12):69−74. DOI: 10.3969/j.issn.1001-8581.2013.12.018 MAO J M, YUE J Q, GAO J Y. Research progress in thrips of fruit trees in China [J]. Acta Agriculturae Jiangxi, 2013, 25(12): 69−74. (in Chinese) DOI: 10.3969/j.issn.1001-8581.2013.12.018
[3] 张小亚, 陈国庆, 黄振东, 等. 为害柑橘的蓟马种类及其防控技术 [J]. 浙江柑橘, 2014, 31(3):27−30. ZHANG X Y, CHEN G Q, HUANG Z D, et al. Species of thrips harming citrus and their control techniques [J]. Zhejiang Ganju, 2014, 31(3): 27−30. (in Chinese)
[4] WEBSTER K W, COOPER P, MOUND L A. Studies on Kelly’s citrus thrips, Pezothrips kellyanus (Bagnall) (Thysanoptera: Thripidae): Sex attractants, host associations and country of origin [J]. Australian Journal of Entomology, 2006, 45(1): 67−74. DOI: 10.1111/j.1440-6055.2006.00508.x
[5] 于法辉, 夏长秀, 方贻文, 等. 引起柑橘果面伤痕的蓟马种类及其发生规律 [J]. 华中农业大学学报, 2014, 33(3):38−41. DOI: 10.3969/j.issn.1000-2421.2014.03.007 YU F H, XIA C X, FANG Y W, et al. Species of thrips causing citrus fruit surface scars and its occurrence regularity [J]. Journal of Huazhong Agricultural University, 2014, 33(3): 38−41. (in Chinese) DOI: 10.3969/j.issn.1000-2421.2014.03.007
[6] VONO G, BONSIGNORE C P, MARULLO R. A comprehensive thrips species assessment for eco-consistent management of infestations in Mediterranean citrus crops [J]. Horticulturae, 2022, 8(2): 137. DOI: 10.3390/horticulturae8020137
[7] 赖跃先. 平和琯溪蜜柚蓟马的发生规律与防治措施 [J]. 福建热作科技, 2018, 43(2):31−33. DOI: 10.3969/j.issn.1006-2327.2018.02.010 LAI Y X. Thrips occurrence rule and preventative measurement on Pinghe Guanxi pomelo [J]. Fujian Science & Technology of Tropical Crops, 2018, 43(2): 31−33. (in Chinese) DOI: 10.3969/j.issn.1006-2327.2018.02.010
[8] 谢永辉, 张宏瑞, 刘佳, 等. 传毒蓟马种类研究进展(缨翅目, 蓟马科) [J]. 应用昆虫学报, 2013, 50(6):1726−1736. DOI: 10.7679/j.issn.2095-1353.2013.236 XIE Y H, ZHANG H R, LIU J, et al. Advances in research on vector thrips species(Thysanoptera, Thripidae) [J]. Chinese Journal of Applied Entomology, 2013, 50(6): 1726−1736. (in Chinese) DOI: 10.7679/j.issn.2095-1353.2013.236
[9] ARPAIA M L, MORSE J G. Citrus thrips Scirtothrips citri (Moulton) (Thys. , Thripidae) scarring and navel orange fruit quality in California[J]. Journal of Applied Entomology, 1991, 111(1/2/3/4/5): 28-32.
[10] 徐淑娟, 张宏瑞, 谢永辉, 等. 柑橘蓟马种类和种群季节动态 [J]. 云南农业大学学报(自然科学), 2012, 27(2):170−175,182. XU S J, ZHANG H R, XIE Y H, et al. Species and seasonal population fluctuation of thrips on citrus [J]. Journal of Yunnan Agricultural University (Natural Science), 2012, 27(2): 170−175,182. (in Chinese)
[11] 秦元霞. 柑橘访花昆虫种类及橘园蓟马的种类、为害、发生规律与防治研究[D]. 武汉: 华中农业大学, 2010. QIN Y X. Study on the species of flower-visiting insects in Citrus and the species, damage, occurrence and control of thrips in citrus orchard[D]. Wuhan: Huazhong Agricultural University, 2010. (in Chinese)
[12] MURAI T. Development and reproductive capacity of Thrips hawaiiensis (Thysanoptera: Thripidae) and its potential as a major pest [J]. Bulletin of Entomological Research, 2001, 91(3): 193−198. DOI: 10.1079/BER200186
[13] MASUI S. Synchronism of immigration of adult yellow tea thrips, Scirtothrips dorsalis Hood (Thysanoptera: Thripidae) to citrus orchards with reference to their occurrence on surrounding host plants [J]. Applied Entomology and Zoology, 2007, 42(4): 517−523. DOI: 10.1303/aez.2007.517
[14] BELAAM-KORT I, MANSOUR R, ATTIA S, et al. Thrips (Thysanoptera: Thripidae) in northern Tunisian Citrus orchards: Population density, damage and insecticide trial for sustainable pest management [J]. Phytoparasitica, 2021, 49(4): 527−538. DOI: 10.1007/s12600-021-00906-y
[15] 姜建军, 黄立飞, 陈红松, 等. 广西4种果树花期主要蓟马种类调查 [J]. 中国南方果树, 2018, 47(2):75−78. JIANG J J, HUANG L F, CHEN H S, et al. Investigation of the species of Thrips at florescence of four fruit plants in Guangxi [J]. South China Fruits, 2018, 47(2): 75−78. (in Chinese)
[16] 丁春霞, 史梦竹, 李建宇, 等. 蓟马类害虫在不同品种芒果上的种群动态及寄主选择性 [J]. 福建农业学报, 2019, 34(8):947−951. DING C X, SHI M Z, LI J Y, et al. Population dynamics of Thrips infesting variety of mango cultivars [J]. Fujian Journal of Agricultural Sciences, 2019, 34(8): 947−951. (in Chinese)
[17] GRAFTON-CARDWELL E E, O’CONNELL N V, KALLSEN C E, et al. Photographic Guide to Citrus Fruit Scarring[M]. University of California, Agriculture and Natural Resources, 2003.
[18] 张宏瑞, Okajim ShÛji, Laurence A. Mound. 蓟马采集和玻片标本的制作 [J]. 昆虫知识, 2006, 43(5):725−728. ZHANG H R, SHÛ JI O, MOUND L. Collecting and slide preparation methods of thrips [J]. Chinese Bulletin of Entomology, 2006, 43(5): 725−728. (in Chinese)
[19] 杨建东, 王自然, 郭俊, 等. 柠檬园蓟马种群动态调查 [J]. 热带农业科学, 2019, 39(3):51−55. YANG J D, WANG Z R, GUO J, et al. Survey of the population dynamics of Thrips in lemon orchards [J]. Chinese Journal of Tropical Agriculture, 2019, 39(3): 51−55. (in Chinese)
[20] 高瑞贺, 范世明, 董江海, 等. 关帝山不同海拔昆虫功能群特征及分布格局 [J]. 生物多样性, 2023, 31(10):109−119. DOI: 10.17520/biods.2023152 GAO R H, FAN S M, DONG J H, et al. Characteristics and vertical distribution of insect functional groups along an altitude gradient in Guandi Mountains [J]. Biodiversity Science, 2023, 31(10): 109−119. (in Chinese) DOI: 10.17520/biods.2023152
[21] RAUT A M, PAL S, WAHENGBAM J, et al. Population dynamics of onion thrips (Thrips tabaci Lindeman, Thysanoptera; Thripidae) and varietal response of onion cultivars against onion thrips [J]. Journal of Entomological Research, 2020, 44(4): 547−554. DOI: 10.5958/0974-4576.2020.00092.4
[22] 武英, 张天翔, 史国强, 等. 琯溪蜜柚蓟马发生为害规律及药剂防效研究 [J]. 中国植保导刊, 2024, 44(1):56−61. DOI: 10.3969/j.issn.1672-6820.2024.01.010 WU Y, ZHANG T X, SHI G Q, et al. Study on the occurrence and control of thrips in Guanxi pomelo in southern Fujian [J]. China Plant Protection, 2024, 44(1): 56−61. (in Chinese) DOI: 10.3969/j.issn.1672-6820.2024.01.010
[23] GRANADA R M, CEBALLO F A, JAVIER P A. Identity and population density of Thrips hawaiiensis (Insecta: Thysanoptera) in conventional "Magallanes"pummelo orchard in Carmen, Davao del Norte, Philippines [J]. The Philippine Agricultural Scientist, 2021, 104(3): 238−247.
[24] KÖRNER C. The use of ‘altitude’ in ecological research [J]. Trends in Ecology & Evolution, 2007, 22(11): 569−574.
[25] ZHAO L J, GAO R H, LIU J Q, et al. Effects of environmental factors on the spatial distribution pattern and diversity of insect communities along altitude gradients in guandi mountain, China [J]. Insects, 2023, 14(3): 224. DOI: 10.3390/insects14030224
[26] FAROOQ M A, ATTA B, ARIF M J, et al. Effect of population dynamics of thrips on blemishes in relation to abiotic factors on Fruiter Early cultivar of citrus [J]. Pakistan Entomologist, 2019, 41(2): 179−186.
[27] 柯胜兵, 党凤花, 毕守东, 等. 不同海拔茶园害虫、天敌种群及其群落结构差异 [J]. 生态学报, 2011, 31(14):4161−4168. KE S B, DANG F H, BI S D, et al. Differences among population quantities and community structures of pests and their natural enemies in tea gardens of different altitudes [J]. Acta Ecologica Sinica, 2011, 31(14): 4161−4168. (in Chinese)
[28] ATAKAN E. Factors affecting fruit damage caused by Thrips hawaiiensis (Morgan) (Thysanoptera: Thripidae) in Citrus lemon groves in Turkey [J]. Phytoparasitica, 2024, 52(1): 25. DOI: 10.1007/s12600-024-01135-9
[29] 李强, 刘奎, 付步礼, 等. 不同食料对黄胸蓟马生物学特性的影响 [J]. 环境昆虫学报, 2018, 40(1):136−143. LI Q, LIU K, FU B L, et al. Effects of different diets on the biological characteristics of the Thrips hawaiiensis [J]. Journal of Environmental Entomology, 2018, 40(1): 136−143. (in Chinese)
[30] HORGAN F G, GARCIA C P F, HAVERKORT F, et al. Changes in insecticide resistance and host range performance of planthoppers artificially selected to feed on resistant rice [J]. Crop Protection, 2020, 127: 104963. DOI: 10.1016/j.cropro.2019.104963
[31] CAO Y, WANG L J, YANG S Y, et al. Population performance of Thrips hawaiiensis (Thysanoptera: Thripidae) on different vegetable host plants [J]. Neotropical Entomology, 2021, 50(6): 923−928. DOI: 10.1007/s13744-021-00914-x
-
期刊类型引用(1)
1. 王玉洁,宋帅,赵艳艳. 青海茄参McCCoAOMT基因的克隆及其在冷胁迫下的表达分析. 青海大学学报. 2025(01): 55-61 .  百度学术
百度学术
其他类型引用(2)

 下载:
下载:











计量
- 文章访问数: 123
- HTML全文浏览量: 9
- PDF下载量: 34
- 被引次数: 3
