

Identification and Genetic Evolution of Viruses Infecting Chili Peppers at Chifeng, Inner Mongolia
-
摘要:目的 系统检测内蒙古赤峰市辣椒主产区病毒发生种类,并分析病毒的遗传分化情况,以期为该地区辣椒病毒病精准防控提供理论依据。方法 从内蒙古赤峰地区4个辣椒主产区采集疑似病毒侵染的辣椒样本47份,采用sRNA高通量测序技术结合生物信息学分析,以及常规RT-PCR检测,明确侵染辣椒的病毒种类及复合侵染情况;采用常规Sanger测序测定常规RT-PCR扩增片段序列,进行遗传进化分析。结果 内蒙古赤峰地区4个辣椒主产区辣椒混合病样的sRNA高通量测序,共检测到6种病毒,常规RT-PCR共验证到其中4种病毒;4个地区病毒种类存在明显差异,松山区城子乡优势种类为蚕豆萎蔫病毒2(Broad bean wilt virus 2,BBWV2)和马铃薯Y病毒(Potato virus Y, PVY),喀喇沁旗锦山镇为PVY和辣椒轻斑驳病毒 (Pepper mild mottle virus, PMMoV);宁城县和喀喇沁旗西桥镇为BBWV2;复合侵染类型以2种病毒复合侵染为主,复合侵染率最高达61.54%。对测定的序列系统发育分析表明,赤峰地区4个辣椒产区的PMMoV和BBWV2存在遗传分化,PVY可能出现了新的遗传型。结论 内蒙古赤峰地区不同辣椒主产区发生的病毒种类、检出率和病毒复合侵染种类均存在明显差异。Abstract:Objective Viruses that infected chili pepper crops in Chifeng City, Inner Mongolia were identified, and their genetic relationship analyzed.Methods The 47 infected chili pepper plants at 4 major producing areas in Chifeng City were diagnosed, and viruses identified by sRNA high-throughput sequencing with bioinformatic analysis and RT-PCR. Fragments of RT-PCR were sequenced using the Sanger method to conduct a phylogenetic analysis with the MEGA software.Results Six viruses were identified by sRNA high-throughput sequencing. Of which, 4 were re-verified by RT-PCR. The dominant viruses infecting the 4 regions differed significantly. They were Broad bean wilt virus 2(BBWV2) and Potato virus Y(PVY) in Chengzi County of Songshang District, PVY and Pepper mild mottle virus(PMMoV) in Jinshan County of Kalaqin District, and BBWV2 in Ningcheng County and Xiqiao County of kalaqin District. When the infection was caused by multiple viruses, it was mostly by two major pathogens and had the highest rate of occurrence at 61.54%. The phylogenetic analysis on the sequenced nucleotides of the viruses showed distinctive genetic differences between PMMoV and BBWV2 and, possibly, PVY of a novel genetic type.Conclusion There are significant differences in the types of viruses, detection rates, and types of virus co-infections that occur in different chili production areas of Chifeng City in Inner Mongolia .
-
0. 引言
【研究意义】松褐天牛Monochamus alternatus Hope,又名松墨天牛,是我国南方松林分布区的重要蛀干害虫,以幼虫钻蛀松树枝干的韧皮部和木质部,影响水分、养分在树体内的传输,致使树体生长衰弱,有的甚至枯死,造成直接的经济损失。同时,松褐天牛也是松材线虫Bursaphelenchus xylophilus Nickle的重要传播媒介。松褐天牛是典型的钻蛀性害虫,除成虫期外,其他虫态均在树干内取食危害,林间天牛种群密度调查及其防治极为困难。【前人研究进展】目前国内学者在松褐天牛的生物学特性、发生规律和防控技术等方面做了大量研究[1-6]。国内有关松褐天牛在马尾松上的垂直分布情况及其与松树胸径、树高的关系也有少量研究报道:孟俊国等研究了松褐天牛幼虫在马尾松上的分布规律[7];高尚坤等系统地研究了松褐天牛产卵刻槽、幼虫和蛹在马尾松树干上的分布规律[8];涂业苟等研究了松褐天牛产卵刻槽、侵入孔和羽化孔的数量与树干胸径的关系[9];张世渊等对浙东沿海地区松褐天牛的成虫羽化历期和各虫态在病树上的分布规律进行了观察研究,同时认为,松褐天牛幼虫及蛹在树体上的分布规律及其保存率是制订清除松材线虫病死树质量标准的主要依据[10]。【本研究切入点】虽然有关学者已对松褐天牛在马尾松树干上的分布规律进行了报道[7-13],但这些研究是某一区域或某一时间段天牛的危害痕迹或幼虫在树干上的分布。天牛在松疫木中取食危害的过程中,会产生产卵刻槽、侵入孔和羽化孔等危害痕迹,危害痕迹调查法是钻蛀性害虫的重要调查方法之一[14]。对于松褐天牛在赣南松材线虫病疫木及其伐桩上的产卵刻槽、侵入孔和羽化孔的数量关系,目前还没有相关报道。【拟解决的关键问题】本研究对松材线虫病死树上的松褐天牛产卵刻槽、侵入孔和羽化孔的垂直分布情况及其数量关系进行了系统的调查分析,在实际调查工作中,调查整株疫木的侵入孔、羽化孔需要花大量人力、物力,针对此情况,本研究探讨疫木下段侵入孔数与整株羽化孔数(虫口密度)的关系,以期探明虫口调查的简便方法,旨在为松褐天牛的林间虫情调查和危害程度估计奠定基础。
1. 材料与方法
1.1 试验地概况
试验地设在江西省赣州市峰山国家森林公园管理处的南田村和东风村,试验林地林分为马尾松Pinus massoniana Lamb.纯林,树龄10~40 a,胸径6~40 cm,平均树高7.5 m,郁闭度0.7~0.9。近年来,该林区出现大量由松材线虫病及松褐天牛造成的马尾松枯死现象,松褐天牛种群密度高,每年均有冬季清理枯死木及综合治理。
1.2 试验方法
每年11月至翌年3月,每月选取10~20 a生、松褐天牛数量多的松材线虫病病死木12株左右,共63株,沿地面伐倒,以株为单位测量树高、胸径,分段统计树干和侧枝上天牛产卵刻槽数,就地放置于林间,待翌年成虫完全羽化后,去皮逐株逐段调查统计幼虫侵入孔和成虫羽化孔的数量。
由于树干长度不一致,在分析时以树干总长度为1,参照杨子祥的方法[14],从基部开始往上1/3处记为下段,往上1/3~2/3处为中段,往上2/3~3/3处为上段,将树干分为下段、中段和上段,再加上侧枝,分段分别统计侵入孔和羽化孔数量,进行分布特征分析。
1.3 试验数据分析
所有数据的统计分析均采用SPSS 17.0统计软件进行方差分析(one-way ANOVA)、t 检验和线性回归分析(Linear regression)。
2. 结果与分析
2.1 松褐天牛虫孔在松材线虫病疫木上的分布
松褐天牛虫孔(产卵刻槽、侵入孔、羽化孔)在马尾松松材线虫病病死树及其伐桩上的分布情况见表 1。
表 1 松褐天牛虫孔在松材线虫病疫木上的分布Table 1. Distribution of M. alternatus boreholes on infected pine trees项目
Item株高
Height/m伐桩
Stumps(5 cm)下段
Lower segment中段
Middle segment上段
Upper segment合计
Total产卵槽数/个
Number of oviposition slots< 8 0 20.55±16.18Bb 34.97±36.51 A b 20.39±11.73B a 75.90±62.41 b ≥ 8 0 81.38±47.97Aa 50.47±22.93ABa 19.75±9.72 B a 151.59±56.24a 侵入孔数/个
Number of intrusive holes< 8 0.66±0.83 a 9.29±6.91 C b 15.84±10.35 A b 11.52±4.10 B a 36.65±19.69 b ≥ 8 0.19±0.40 b 35.03±19.77Ba 49.88±34.27 A a 18.88±13.86C a 103.81±58.62a 羽化孔数/个
Number of eclosion holes< 8 0.29±0.46 a 5.45±3.71 Cb 9.39±4.96 A b 7.25±3.03 B b 22.10±10.27 b ≥ 8 0.10±0.30 b 18.00±9.75 Ba 26.31±16.85 A a 11.03±7.57 BCa 55.34±28.02 a 注:表中同一行不同大写字母表示差异显著(P < 0.05),相同项目下的每一列不同小写字母表示差异显著(P < 0.05);表中数字为“平均值±标准差”。
Note:Different capital letters on a same row indicate significant differences (P < 0.05). Different lowercase letters on a same column indicate significant differences (P < 0.05). Datas are mean±SD.由表 1可见,所调查的整株松褐天牛产卵刻槽数量介于25~259个,平均114.35个。株高 < 8 m的植株平均刻槽数为75.90个,其中下段20.55个、中段34.97个、上段20.39个,分别占总刻槽数的27.07%、46.07%、26.86%,产卵刻槽在病树上的数量分布特征为:中段>下段>上段,其中中段占比近50%,与上、下段的差异均达极显著水平(P < 0.01),而后两者差异不显著(P>0.05);株高≥ 8 m的植株平均刻槽数为151.59个,其中下段81.38个、中段50.47个、上段19.75个,分别占总刻槽数的53.687%、33.29%、13.03%,产卵刻槽在病树上的数量分布特征为:下段>中段>上段,其中下段占比超50%,与上段差异达极显著水平(P < 0.01)。可见,株高 < 8 m的整株产卵刻槽主要分布在树干中段,而株高≥ 8 m的则主要分布在树干下段,回归分析结果显示,松褐天牛产卵刻槽数量与株高的关系呈显著正相关关系(R2=0.479; df=1, 62; F=56.147; t=7.493; P < 0.01)。
所调查的整株天牛侵入孔数量介于17~221个,平均70.76个,其中下段22.37个、占总数的33.76%,中段33.13个、占48.04%,上段15.25个、占18.21%,在病树上的数量分布特征为:中段>下段>上段,且中段数量显著大于下、上段数量。回归分析结果显示,松褐天牛侵入孔数量与株高的关系呈显著正相关关系(R2=0.418; df=1, 62; F=43.794; t=6.618; P= < 0.01),不管株高大于还是小于8 m,均为中段的侵入孔数量最多。
所调查的整株天牛羽化孔数量介于9~109个,平均38.98个,其中下段11.83个、占总数的32.52%,中段17.98个、占47.54%,上段9.18个、占19.93%,在病树上的数量分布特征为:中段>下段>上段,中段羽化孔数量最多,均占近50%,与上、下段的差异均达极显著水平(P < 0.01)。回归分析结果显示,松褐天牛羽化孔数量与株高的关系呈显著正相关关系(R2=0.353; df=1, 62; F=33.216; t=5.763; P < 0.01)。
2.2 产卵刻槽、侵入孔和羽化孔三者间的关系分析
病死树上松褐天牛的产卵刻槽、侵入孔和羽化孔三者间的关系见表 2。由表 2可见,株高 < 8 m的植株上,天牛产卵刻槽数:侵入孔数:羽化孔数=1:0.483:0.291;株高≥ 8 m的植株上,天牛产卵刻槽数:侵入孔数:羽化孔数=1:0.685:0.365。相关分析结果显示:所有P < 0.01,表明两两间存在明显的相关关系,所有的相关系数R≥0.720,两两间为极显著相关,回归方程见表 2。
表 2 产卵刻槽、侵入孔和羽化孔数量间的关系分析Table 2. Correlations among numbers of oviposition grooves, invasion and eclosion holes on diseased tree trunks株高
Height/m项目
Item比例
Proportion相关系数
RP值
Sig.回归方程
Regression equation< 8 产卵刻槽数x1与侵入孔数x2
No of oviposition scars (x1) and No of entrance holes (x2)1:0.483 0.918 0.000 x2=0.290x1 + 14.665 产卵刻槽数x1与羽化孔数x3
No of oviposition scars (x1) and No of exit holes (x3)1:0.291 0.720 0.000 x3=0.119x1 + 13.100 侵入孔数x2与羽化孔数x3
No of entrance holes (x2) and No of exit holes (x3)1:0.603 0.911 0.000 x3=0.475x2 + 4.677 ≥ 8 产卵刻槽数x4与侵入孔数x5
No of oviposition scars (x4) and No of entrance holes (x5)1:0.685 0.848 0.000 x5=0.884x4-30.263 产卵刻槽数x4与羽化孔数x6
No of oviposition scars (x4) and No of exit holes (x6)1:0.365 0.721 0.000 x6=0.359x4 + 0.863 侵入孔数x5与羽化孔数x6
No of entrance holes (x5) and No of exit holes (x6)1:0.533 0.950 0.000 x6=0.454x5 + 8.207 2.3 下段虫孔数量与整株羽化孔数量的关系
马尾松疫木下段虫孔数量(即下段产卵刻槽、侵入孔和羽化孔数量)与整株羽化孔数量的相关分析表明:下段产卵刻槽、侵入孔和羽化孔的数量均与整株羽化孔数量存在明显的相关关系(表 3),所有相关系数≥ 0.834,P < 0.01,分别达极显著相关,其回归方程见表 3。这一结果表明,松材线虫病疫木虫口密度调查,可以依据下段产卵刻槽、侵入孔或羽化孔的数量来估计整株羽化孔数量,从而推算林间的天牛虫口数量,制定合理的防治方案。
表 3 下段虫孔数量与整株羽化孔数量相关性的回归分析Table 3. Regression analysis for correlation between numbers of boreholes on lower section and total eclosion holes on a diseased-tree trunk项目
Item相关系数
R决定系数
R2F值
Ft值
tP值
Sig.回归方程
Regression equation整株羽化孔数y
No of emergent holes in the whole plant下段刻槽数x1
No of grooves in lower section (x1)0.834 0.695 139.118 11.795 0.000 y=0.477x1+14.4675 下段侵入孔数x2
No of entrance holes in lower section (x2)0.928 0.861 377.493 19.429 0.000 y=1.270x2+10.588 下段羽化孔数x3
No of exit holes in lower section (x3)0.941 0.886 473.915 21.770 0.000 y=2.610x3+8.116 2.4 松褐天牛虫孔在松病死木伐桩上的分布
病死木伐桩地上部松褐天牛产卵刻槽、侵入孔及羽化孔情况见表 1。从表 1可见,调查的病死木伐桩上未发现有天牛产卵刻槽,侵入孔和羽化孔数量也极少。病死木伐桩地上部天牛侵入孔平均每株0.48个,其中株高 < 8 m的疫木其伐桩侵入孔平均数量每桩为0.66个,株高≥ 8 m的为0.19个,两者差异达显著水平(P < 0.05);病死木伐桩地上部天牛羽化孔平均每株0.14个,其中株高 < 8 m的疫木其伐桩羽化孔平均数量每桩为0.29个,株高≥ 8 m的为0.10个,两者差异达显著水平(P < 0.05)。
可见,病死树越高,伐桩上虫孔数量越少,但相关性不显著(R2 =0.082),因为每株伐桩虫孔数极少,在0~3个。有虫孔株率分别为38.10%、14.29%,病死木伐桩地上部天牛侵入孔数、羽化孔数分别占总株侵入孔数、羽化孔数的0.54%、0.29%,表明相对于全株上天牛种群数量而言,病死木伐桩上虽有天牛存在,但数量极低,即松褐天牛成虫的出现率较低,有羽化成虫的有虫伐桩率为14.29%,伐桩羽化成虫数量占整株羽化成虫数的0.35%。
3. 讨论与结论
本研究发现,松材线虫病病死树上的松褐天牛产卵刻槽的数量分布特征,株高 < 8 m的植株,整株产卵刻槽主要分布在树干中段,占46.07%,而株高≥8 m的植株,整株产卵刻槽主要分布在树干下段,占53.687%,其次是中段,产卵刻槽数量与株高的关系均呈显著正相关关系。这与涂业苟等[9]、杨子祥等[14]的研究结论相一致:松褐天牛产卵刻槽的数量主要集中分布在马尾松树干1~5 m的位置,树干下段分布最多。这与高娜等[15]的研究不一致:黑松树干3 m以上很少有天牛产卵刻槽分布,树高>7 m时,没有产卵刻槽分布。出现这些差异的原因可能与松树品种、松树生长状况(包括树皮厚度、树干含水量和树皮内营养成分)有关[14-15],有待进一步研究。
本试验显示,松褐天牛侵入孔、羽化孔在被害病死树上的数量分布特征为树干中段>下段>上段,且中段数量显著大于下段、上段数量,这与前人的研究结论一致[10, 12, 14]。天牛产卵刻槽、侵入孔和羽化孔,两两间存在明显的数量相关关系且显著相关。根据王玲萍[16]的研究:松褐天牛在马尾松树干内卵的孵化率为86.18%, 孵化后的幼虫只有31.68%能进入木质部,各段的孵化率或侵入率的差异可能是造成侵入孔与产卵刻槽分布特征不一致的原因。而幼虫蛀入木质部后,受环境因素的影响较小,生存率较高[10],因此羽化孔和侵入孔出现了相似的分布特征,即中段分布最多,全树幼虫侵入孔与成虫羽化孔的数量比例稳定在1:0.5~1:0.6。
关于松褐天牛的林间虫情调查问题。松褐天牛是典型的钻蛀性害虫,危害痕迹(产卵刻槽、侵入孔和羽化孔)调查法是钻蛀性害虫的重要调查方法之一[14]。在实际调查工作中,调查整株疫木的侵入孔、羽化孔需要花大量人力、物力,难度很大,而树干下段的危害痕迹比较容易调查,根据下段危害痕迹数量来估计整株的羽化孔数量,有助于快速掌握林间的虫口数量[14]。本研究结果显示,树干下段虫孔数量(即下段产卵刻槽、侵入孔和羽化孔数量)与整株羽化孔数量呈极显著相关关系,即均可采用下段产卵刻槽、侵入孔或羽化孔数量来估计整株的羽化孔数量。有报道认为:根据整株(或树干高度2~3 m)刻槽的数量,可以预测整株马尾松上的松褐天牛羽化孔总数量,更直观可靠[9]。但由于无效产卵刻槽的存在[4, 16-17],采用产卵刻槽来估计羽化孔数量,可能会产生较大的误差[14]。松褐天牛成虫羽化历期太长(4~8月),且松材线虫病病死树的清除一般须在4月前完成,所以采用下段羽化孔数量来估计整株羽化孔数量也是不现实的。而天牛幼虫则较早进入木质部,蛀入后受环境条件和天敌影响较小,种群数量基本稳定[10],因此根据下段侵入孔数量来推算整株羽化孔的数量更为准确、可靠。
本研究还发现,在所调查的松材线虫病病死树中,伐桩上没有发现产卵刻槽,有松褐天牛侵入孔、羽化孔的伐桩数分别占总伐桩数的38.10%、14.29%,伐桩内虽有松褐天牛分布,但数量极少,每桩虫孔数量在0~3个,伐桩羽化成虫数量占整株羽化成虫数的0.35%。而伐桩暴露在林间的时间长,伐桩内的天牛一般在5~6月羽化,因此,虽然有的伐桩上可能有松褐天牛成虫羽化,但其虫体也可能不带有松材线虫(另文发表),即不能发挥媒介作用,这与蒋丽雅等[18]的研究结论一致。因此,我们认为,在松材线虫病疫木清理过程中,只要控制好伐桩高度(低于5 m),伐桩则无需处理。
-
![]()
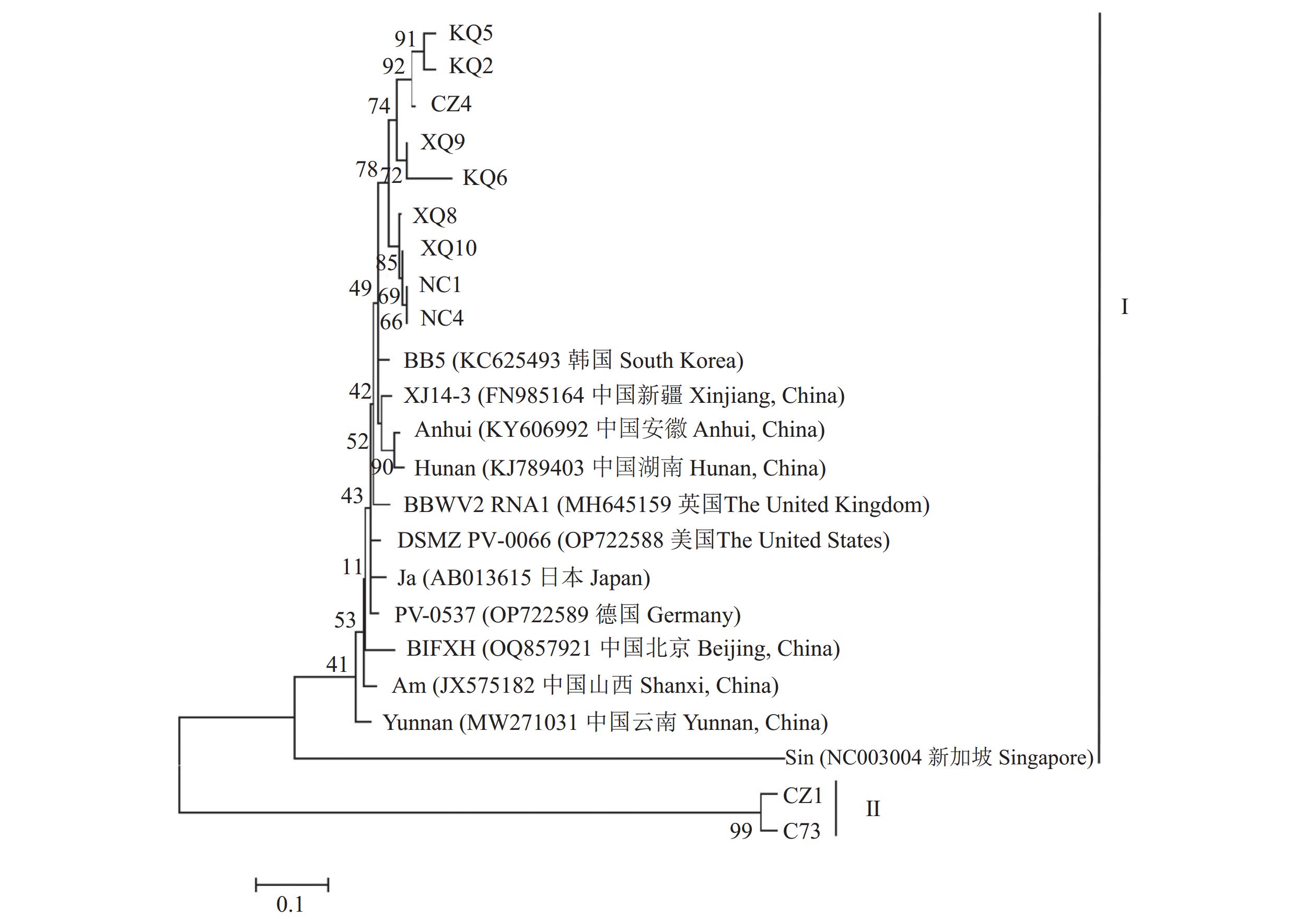
图 2 BBWV2 RNA依赖的RNA聚合酶基因部分核苷酸序列系统发育分析
Figure 2. Phylogenetic analysis on partial nucleotide sequence of RNA dependent RNA polymerase gene of BBWV2
![]()
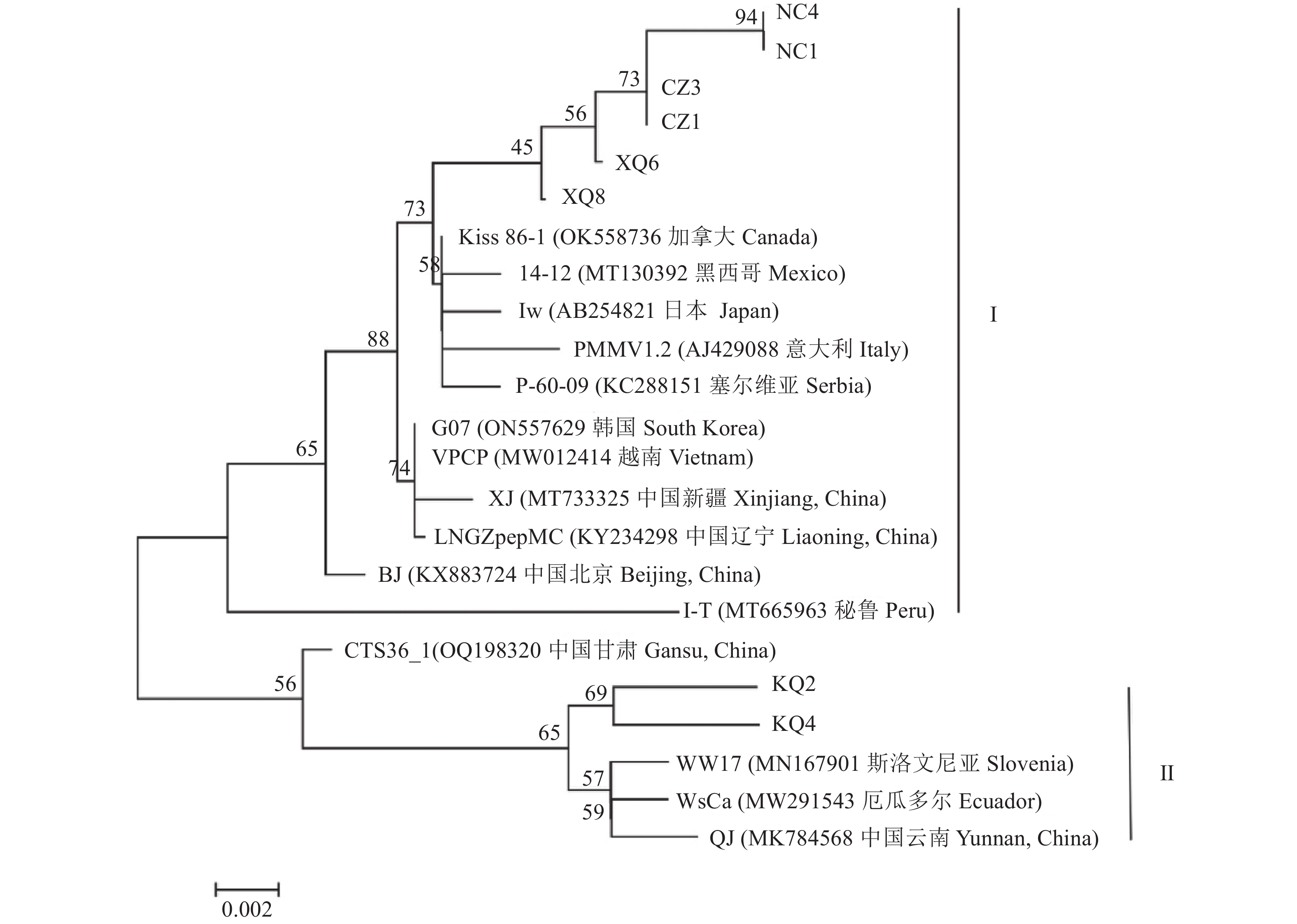
图 3 PMMoV CP基因部分核苷酸序列系统发育分析
Figure 3. Phylogenetic analysis on partial nucleotide sequence of CP gene of PMMoV
![]()
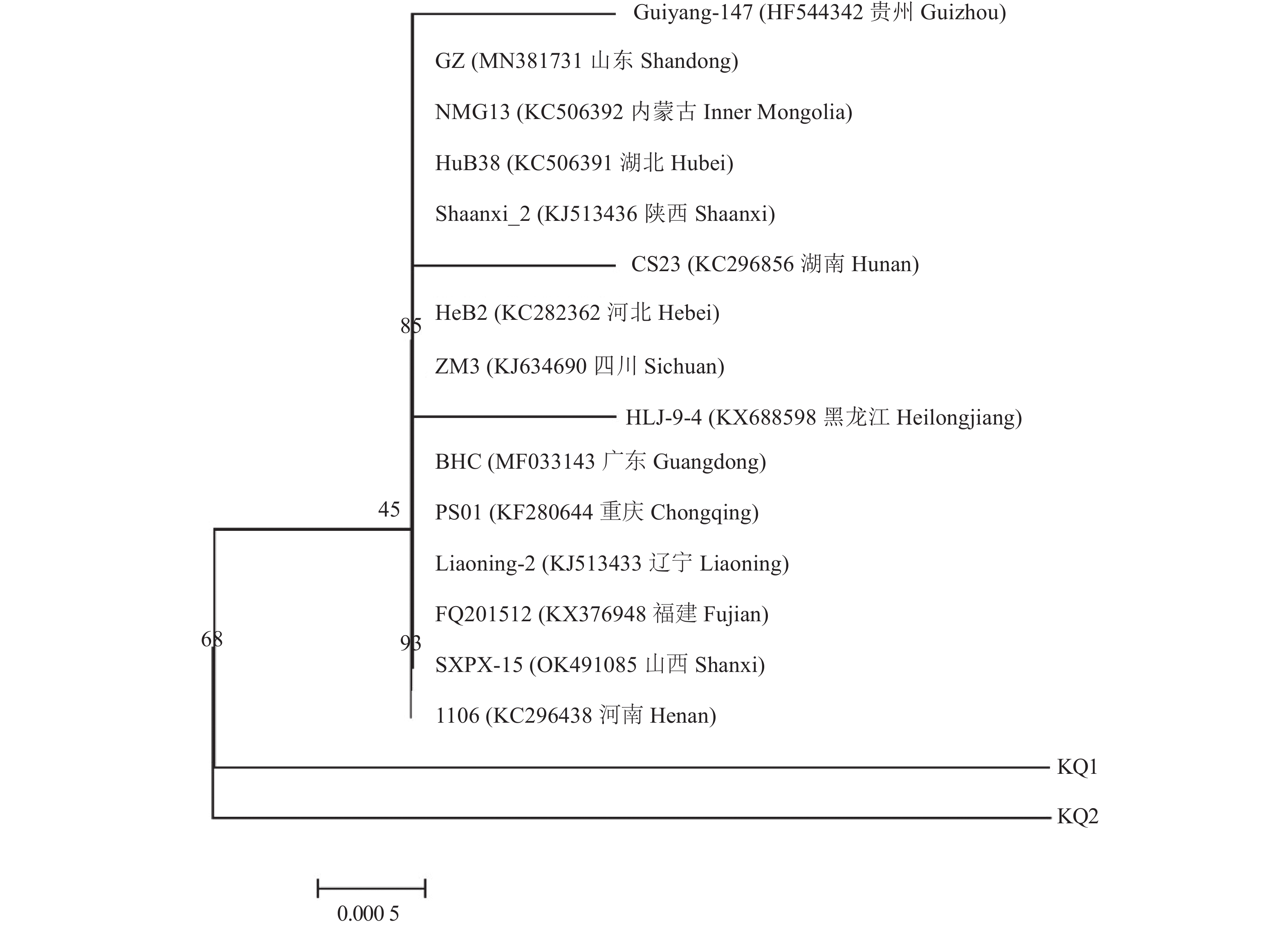
图 4 PVY CP基因部分核苷酸序列系统发育分析
Figure 4. Phylogenetic analysis on partial nucleotide sequence of CP gene of PVY
表 1 sRNA测序数据质量
Table 1 Quality of data obtained by sRNA
样本
编号
Sample No.原始
读数量
Reads/M原始
序列量
Bases /MGC含量
GC content/
%质量值Q30/% 片段数量
Contig
number片段长
度中值
Contig N50/bp城子乡CZ 12.14 607 52.34 97.45 284 157 锦山镇KQ 13.34 667 48.28 97.46 394 182 宁城县NC 13.43 671 51.06 97.37 276 168 西桥镇XQ 12.80 640 51.64 97.60 258 146 Q30:Phred 数值大于 30 的碱基占总体碱基的百分比。
Q30: ratio of bases of Phred number over 30 in total bases. 下载: 导出CSV
下载: 导出CSV
表 2 辣椒病毒检出率
Table 2 Positive viral detection rate on chili pepper plants
采样地点
Sampling site辣椒样本病毒检出率
Rates of positive detection of viruses in pepper plants/%SPVG PMMoV BBWV2 PVY INLV SCMV 城子乡CZ 0 61.54 92.31 0 23.08 0 锦山镇KQ 0 58.33 41.67 83.33 0 0 宁城县NC 0 25.00 58.33 0 0 0 西桥镇XQ 0 20.00 30.00 0 0 0
下载: 导出CSV
表 3 辣椒病毒复合侵染率
Table 3 Occurrence rate of multiple viruses infection on chili pepper plants
采样地点
Sampling site病毒复合侵染种类
Viruses of fused infection病毒检出率
Rates of positive
detection of viruses/%城子乡CZ BBWV2+PMMoV 61.54 BBWV2+INLV 23.08 BBWV2+INLV+PMMoV 23.08 锦山镇KQ PMMoV+BBWV2 41.67 PMMoV+PVY 58.33 BBWV2+PVY 41.67 PMMoV+BBWV2+PVY 25.00 宁城县NC PMMoV+BBWV2 25.00 西桥镇XQ PMMoV+BBWV2 15.38
下载: 导出CSV
-
[1] 崔聪聪, 王秀芝, 张晓梅, 等. 赤峰市露地辣椒生产情况及经济效益调查分析 [J]. 现代农业科技, 2018, (24):92−93,95. DOI: 10.3969/j.issn.1007-5739.2018.24.054 CUI C C, WANG X Z, ZHANG X M, et al. Investigation and analysis on the production situation and economic benefit of pepper in Chifeng city [J]. Modern Agricultural Science and Technology, 2018(24): 92−93,95. (in Chinese) DOI: 10.3969/j.issn.1007-5739.2018.24.054
[2] 张晓梅, 王秀芝, 崔聪聪, 等. 2018年赤峰地区设施辣椒生产与市场情况分析 [J]. 中国蔬菜, 2019, (3):89−92. ZHANG X M, WANG X Z, CUI C C, et al. Production and market analysis of protected pepper in Chifeng area in 2018 [J]. China Vegetables, 2019(3): 89−92. (in Chinese)
[3] 柴阿丽, 陈利达, 曹金强, 等. 利用siRNA高通量测序和RT-PCR技术鉴定引起茄子斑驳紫花病的病毒种类 [J]. 园艺学报, 2019, 46(3):508−518. CHAI A L, CHEN L D, CAO J Q, et al. Identification of viruses causing eggplant purple mottle flower disease by sirna high-throughput sequencing and RT-PCR detection [J]. Acta Horticulturae Sinica, 2019, 46(3): 508−518. (in Chinese)
[4] 刘勇, 李凡, 李月月, 等. 侵染我国主要蔬菜作物的病毒种类、分布与发生趋势 [J]. 中国农业科学, 2019, 52(2):239−261. DOI: 10.3864/j.issn.0578-1752.2019.02.005 LIU Y, LI F, LI Y Y, et al. Identification, distribution and occurrence of viruses in the main vegetables of China [J]. Chinese Agricultural Sciences, 2019, 52(2): 239−261. (in Chinese) DOI: 10.3864/j.issn.0578-1752.2019.02.005
[5] SKELTON A, UZAYISENGA B, FOWKES A, et al. First report of Pepper veinal mottle virus, pepper yellows virus and a novel enamovirus in chilli pepper (Capsicum sp. ) in Rwanda [J]. New Disease Reports, 2018, 37(1): 5. DOI: 10.5197/j.2044-0588.2018.037.005
[6] 冯耿, 辛敏, 曹孟籍, 等. 深度测序发现贵阳发生的辣椒病毒病由多种病毒复合侵染所致 [J]. 植物病理学报, 2017, 47(5):591−597. FENG G, XIN M, CAO M J, et al. Identification of multiple viruses infecting hot pepper in Guiyang by deep sequencing [J]. Acta Phytopathologica Sinica, 2017, 47(5): 591−597. (in Chinese)
[7] 廖震, 赵德刚, 赵懿琛. 利用小RNA深度测序技术检测分析小黄姜病毒 [J]. 基因组学与应用生物学, 2018, 37(6):2417−2422. LIAO Z, ZHAO D G, ZHAO Y C. Detection and analysis of viruses from small yellow ginger (Zingiber officinale Rosc. ) by small RNA deep sequencing technology [J]. Genomics and Applied Biology, 2018, 37(6): 2417−2422. (in Chinese)
[8] 汤亚飞, 裴凡, 李正刚, 等. 基于小RNA深度测序技术鉴定侵染广东辣椒的病毒种类 [J]. 中国农业科学, 2019, 52(13):2256−2267. DOI: 10.3864/j.issn.0578-1752.2019.13.006 TANG Y F, PEI F, LI Z G, et al. Identification of viruses infecting peppers in Guangdong by small RNA deep sequencing [J]. Scientia Agricultura Sinica, 2019, 52(13): 2256−2267. (in Chinese) DOI: 10.3864/j.issn.0578-1752.2019.13.006
[9] 于海龙, 靳远, 刘婧, 等. 我国辣椒病毒病发生情况及发展趋势: 基于2018年和2019年辣椒主产区的调查 [J]. 中国蔬菜, 2020, (9):25−30. YU H L, JIN Y, LIU J, et al. Occurrence and development trend of pepper virus disease in China—based on main pepper producing areas investigation in 2018 and 2019 [J]. China Vegetables, 2020(9): 25−30. (in Chinese)
[10] LANGMEAD B, TRAPNELL C, POP M, et al. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome [J]. Genome Biology, 2009, 10(3): R25. DOI: 10.1186/gb-2009-10-3-r25
[11] TAMURA K, STECHER G, KUMAR S. MEGA11: Molecular evolutionary genetics analysis version 11 [J]. Molecular Biology and Evolution, 2021, 38(7): 3022−3027. DOI: 10.1093/molbev/msab120
[12] VISSER M, BESTER R, BURGER J T, et al. Next-generation sequencing for virus detection: covering all the bases [J]. Virology Journal, 2016, 13: 85. DOI: 10.1186/s12985-016-0539-x
[13] SANTALA J, VALKONEN J P T. Sensitivity of small RNA-based detection of plant viruses [J]. Frontiers in Microbiology, 2018, 9: 939. DOI: 10.3389/fmicb.2018.00939
[14] GARCÍA-ARENAL F, FRAILE A, MALPICA J M. Variability and genetic structure of plant virus populations [J]. Annual Review of Phytopathology, 2001, 39: 157−186. DOI: 10.1146/annurev.phyto.39.1.157
[15] 李桑桑, 胡荣, 罗香文, 等. 湖北和广西辣椒脉斑驳病毒的检测及 遗传多样性分析 [J]. 南方农业学报, 2020, 51(7):1693−1698. DOI: 10.3969/j.issn.2095-1191.2020.07.023 LI S S, HU R, LUO X W, et al. Detection and genetic identity of Pepper veinal mottle virus in Hubei and Guangxi [J]. Journal of Southern Agriculture, 2020, 51(7): 1693−1698. (in Chinese) DOI: 10.3969/j.issn.2095-1191.2020.07.023
[16] 汤亚飞, 裴凡, 于琳, 等. 侵染广东辣椒的辣椒脉斑驳病毒的分子特征 [J]. 园艺学报, 2018, 45(11):2209−2216. TANG Y F, PEI F, YU L, et al. Molecular characterization of Chilli veinal mottle virus infecting pepper in Guangdong province [J]. Acta Horticulturae Sinica, 2018, 45(11): 2209−2216. (in Chinese)
[17] PICARD C, DALLOT S, BRUNKER K, et al. Exploiting genetic information to trace plant virus dispersal in landscapes [J]. Annual Review of Phytopathology, 2017, 55: 139−160. DOI: 10.1146/annurev-phyto-080516-035616
[18] VASSILAKOS N, SIMON V, TZIMA A, et al. Genetic determinism and evolutionary reconstruction of a host jump in a plant virus [J]. Molecular Biology and Evolution, 2016, 33(2): 541−553. DOI: 10.1093/molbev/msv222
[19] FERRER R M, FERRIOL I, MORENO P, et al. Genetic variation and evolutionary analysis of broad bean wilt virus 2 [J]. Archives of Virology, 2011, 156(8): 1445−1450. DOI: 10.1007/s00705-011-0990-3
[20] GUAN X Y, YANG C X, FU J J, et al. Rapid evolutionary dynamics of pepper mild mottle virus [J]. Virus Research, 2018, 256: 96−99. DOI: 10.1016/j.virusres.2018.08.006
[21] GAO F L, ZOU W C, XIE L H, et al. Adaptive evolution and demographic history contribute to the divergent population genetic structure of Potato virus Y between China and Japan [J]. Evolutionary Applications, 2017, 10(4): 379−390. DOI: 10.1111/eva.12459
[22] 龚明霞, 赵虎, 王萌, 等. 广西辣椒病毒病调查及病原种类初步鉴定 [J]. 中国蔬菜, 2020, (4):74−79. GONG M X, ZHAO H, WANG M, et al. Investigation and preliminary identification of pathogeny species of pepper virus disease in Guangxi [J]. China Vegetables, 2020(4): 74−79. (in Chinese)
[23] 郭思瑶, 童艳, 黄娅, 等. 重庆辣椒病毒病病原初步鉴定和分析 [J]. 园艺学报, 2015, 42(2):263−270. GUO S Y, TONG Y, HUANG Y, et al. Preliminary identification and analyses of viruses causing pepper virus disease in Chongqing, China [J]. Acta Horticulturae Sinica, 2015, 42(2): 263−270. (in Chinese)
[24] 王少立, 谭玮萍, 杨园园, 等. 山东省辣椒主要病毒种类的分子检测与鉴定 [J]. 中国农业科学, 2017, 50(14):2728−2738. DOI: 10.3864/j.issn.0578-1752.2017.14.009 WANG S L, TAN W P, YANG Y Y, et al. Molecular detection and identification of main viruses on pepper in Shandong Province [J]. Scientia Agricultura Sinica, 2017, 50(14): 2728−2738. (in Chinese) DOI: 10.3864/j.issn.0578-1752.2017.14.009
[25] 姚玉荣, 陈国华, 冯兰香, 等. 北运蔬菜基地辣椒病毒病病原种类的分子检测 [J]. 中国蔬菜, 2013, (10):84−89. DOI: 10.3969/j.issn.1000-6346.2013.10.014 YAO Y R, CHEN G H, FENG L X, et al. Molecular detection of pepper viruses in southern vegetable production bases [J]. China Vegetables, 2013(10): 84−89. (in Chinese) DOI: 10.3969/j.issn.1000-6346.2013.10.014
[26] PEÑAFLOR M F G V, MAUCK K E, ALVES K J, et al. Effects of single and mixed infections of Bean pod mottle virus and Soybean mosaic virus on host-plant chemistry and host–vector interactions [J]. Functional Ecology, 2016, 30(10): 1648−1659. DOI: 10.1111/1365-2435.12649
[27] LAMICHHANE J R, VENTURI V. Synergisms between microbial pathogens in plant disease complexes: A growing trend [J]. Frontiers in Plant Science, 2015, 6: 385.
[28] SAFARI M, ROOSSINCK M J. Coevolution of a persistent plant virus and its pepper hosts [J]. Molecular Plant-Microbe Interactions, 2018, 31(7): 766−776. DOI: 10.1094/MPMI-12-17-0312-R
-
期刊类型引用(13)
1. 何子婷,张育华,钟先龙,徐雪丽,胡平. 不同松材线虫病感病时期马尾松钻蛀性昆虫差异分析. 广西林业科学. 2025(01): 59-65 .  百度学术
百度学术
2. 王志华,董立坤,毛润萍,于静亚,刘超,段庆明,潘婷婷. 星天牛在悬铃木上的空间分布研究. 林业与环境科学. 2023(02): 116-120 . 百度学术
3. 陈元生,周嘉颖,肖小华,罗致迪,彭小文. 林间花斑花绒寄甲幼虫种群动态及空间分布格局. 环境昆虫学报. 2023(03): 688-694 . 百度学术
4. 薛万全,张丽萍,缪玮. 镇江市绿化植物主要天牛种类及其成虫发生动态. 上海农业科技. 2023(04): 145-146 . 百度学术
5. 陈珍珍. 基于诱捕法的惠安县松褐天牛种群动态研究. 武夷科学. 2022(02): 121-125 . 百度学术
6. 李云松. 昆明市海口林场松材线虫病调查报告. 绿色科技. 2021(01): 136-137 . 百度学术
7. 沈晓旭. 松材线虫病病死木数量回归预测研究. 南方农业. 2021(05): 131-132 . 百度学术
8. 余海. 基于松褐天牛眼高刻槽数预测松材线虫病发生量试验. 现代农业科技. 2021(10): 84-87 . 百度学术
9. 褚东花,李德峰,宋西强. 基于多光谱遥感的松材线虫病受害木识别方法. 绿色科技. 2021(09): 178-180 . 百度学术
10. 陈元生,于海萍,罗致迪. 松材线虫在疫木内的分布动态. 西北农业学报. 2021(03): 462-467 . 百度学术
11. 肖正利,敬顺华,刘海玲,周勇,陈玉萍. 基于GIS空间统计诱捕法的松褐天牛种群动态和空间密度分布测定——以湖北省远安县为例. 林业调查规划. 2021(04): 54-60 . 百度学术
12. 杨丽元,刘仁军,赵筱菲,黄实,华杰,孙守慧. 油松蛀干害虫及其寄生蜂在树干上的垂直分布. 中国生物防治学报. 2021(04): 701-708 . 百度学术
13. 南俊科,杨越翔,张玲华,李鹏飞,郭丽洁,贺虹,魏琮. 松墨天牛在秦巴林区不同寄主上的危害规律. 环境昆虫学报. 2021(06): 1376-1388 . 百度学术
其他类型引用(4)
计量
- 文章访问数: 364
- HTML全文浏览量: 76
- PDF下载量: 52
- 被引次数: 17
