

Response Surface Optimization on Formulation of Cookie Made with Ultramicro Green Tea Powder
-
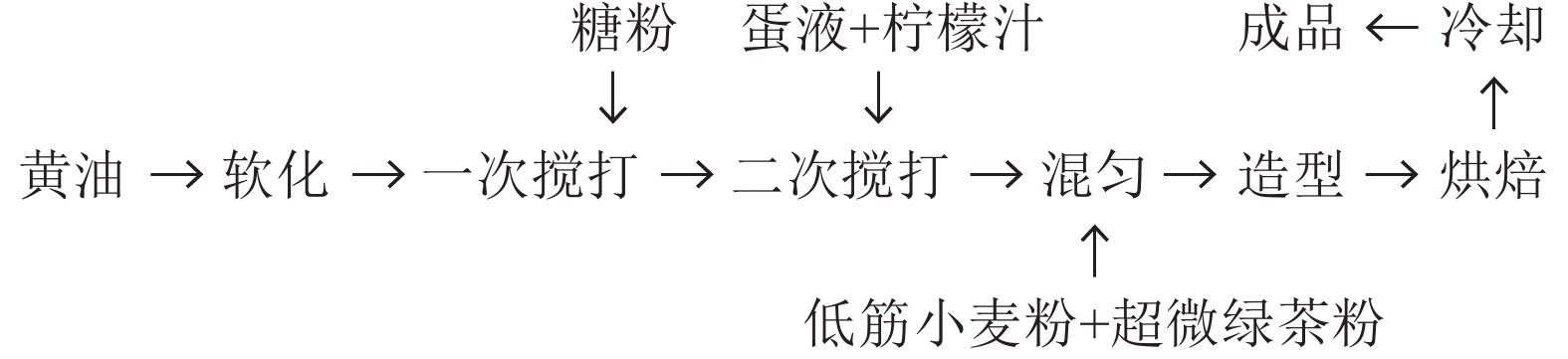
摘要:目的 优化超微绿茶粉曲奇饼干配方,为茶叶精深加工开发终端产品提供思路。方法 对超微绿茶粉、糖粉、黄油的添加量进行单因素试验,通过感官审评对其进行质量评价,确定3个因素添加量的优化范围。在单因素试验基础上,采用Box-Behnken试验设计,进行3因素3水平响应面优化设计,共17组处理,考察超微绿茶粉、糖粉、黄油的添加量对超微绿茶粉曲奇饼干感官审评的影响,确定超微绿茶粉曲奇饼干的配方,并对优化配方加工的饼干进行感官评价、理化指标检测、质构评价。结果 超微绿茶粉制作曲奇饼干的配方为:以总粉(低筋小麦粉+超微绿茶粉)100 g即100%计,蛋液添加量15%,柠檬汁1%,超微绿茶粉3.0%,黄油84.4%,糖粉25.9%,烘焙上火温度170 ℃,下火温度150 ℃,时间20 min。在此条件下加工的超微绿茶粉曲奇饼干,感官审评得分85.36分,风味较好。经质构仪分析,饼干的硬度为578.44 g,酥性为626.46 g·s。经检测,饼干的各项理化指标均符合国标要求。结论 采用响应面法得出了超微绿茶粉曲奇饼干最优的关键配方,可用于规模化生产,为茶叶精深加工开发茶食品、缓解茶产业供需矛盾提供新的思路。Abstract:Objective A formula for making cookies using ultramicro green tea powder as an ingredient was optimized.Method A single factor experiment was conducted to define the recipe including the ultramicro green tea powder, butter, and powdered sugar for making cookies evaluated by a sensory panel. A 3-factors Box-Behnken test was designed to optimize the amount of each ingredient based on the results of sensory evaluation, and the physical, and chemical properties of the cookies with the optimum crecipe were deteceted.Result The cookies baked in an air oven with an overhead temperature of 170 ℃ and a underneath temperature of 150 ℃ for 20 min, received a high sensory evaluation score of 85.36 with 100% of total flour (low-gluten wheat flour + ultramicro green tea powder), 15% of the additive amount of egg liquid, 1% of lemon juice, 3.0% of ultramicro green tea powder, 84.4% of butter, and 25.9% of powdered sugar. Measured by a texture analyzer, the cookies had the indices in line with the national standards on hardness of 578.44 g and on crispness of 626.46 g·s.Conclusion The cookie made with an optimized formula was well received by the sensory panel with acceptable instrumental texture measurements. It was considered suitable for mass producing a value-added product of green teas.
-
Keywords:
- Ultramicro green tea powder /
- cookies /
- response surface method /
- formulation
-
植物病原菌在侵染植物的过程中会分泌一些效应因子(又称效应蛋白,effector)到寄主细胞,随后效应因子靶标到不同寄主蛋白,破坏寄主抗病信号途径进而促进致病[1]。因此,效应因子是病原物毒性水平和寄主范围的关键决定因子[2]。病原物效应因子可抑制病原物相关分子模式(pathogen-associated molecular patter,PAMP)触发的寄主免疫反应(PAMP-triggered immunity,PTI)或效应因子触发的免疫反应(effector-triggered immunity,ETI)[3]。PTI和ETI的反应强度不同,ETI会导致强烈的过敏反应(Hypersensitive Response,HR),这种反应发生在病原物侵染部位并阻止病原物进一步扩散[4-5]。
CFEM(Common in Fungal Extracellular Membrane)是真菌中特有的一类位于细胞外膜的蛋白,许多CFEM结构蛋白被证明在病原真菌与寄主植物互作中起效应因子的作用[6-7]。目前有关真菌CFEM蛋白生物学功能的研究相对匮乏,本文主要综述真菌CFEM蛋白的结构、表达模式与定位、起源和进化过程以及调控真菌铁吸收、生长发育与致病性的功能与机制,对阐明植物-病原真菌互作分子机理、揭示真菌CFEM蛋白的功能奠定基础。
1. CFEM蛋白家族
1.1 CFEM蛋白的结构
CFEM是真菌中特异的常见于细胞外膜的一种蛋白,其结构域约有60个氨基酸,共有8个间隔排列的高度保守的半胱氨酸(图1),其特点是将其与已知的富含半胱氨酸的结构域区分开来。与其他富含半胱氨酸的蛋白不同,该蛋白结构域仅存在于真菌中,是常见的真菌胞外膜蛋白,并因此而得名[6]。CFEM类效应因子可以附着在真菌细胞壁上,并可以转移到糖基磷脂酰肌醇(Glycosylphosphatidylinositol,GPI)锚定位点,在真菌致病过程中发挥重要作用[8]。
自首次从粗球孢子菌(Coccidioides immitis)中鉴定出CFEM类蛋白以来,已经从多种真菌中鉴定出CFEM蛋白,包括白色念珠菌(Candida albicans)、近平滑假丝酵母(Candida parapsilosis)、稻瘟菌(Magnaporthe oryzae)、少孢节丛孢菌(Arthrobotrys oligospora)、禾谷炭疽菌(Colletotrichum graminicola)和烟曲霉(Aspergillus fumigatus)等[6,8-10]。例如,稻瘟菌中发现了19个CFEM蛋白[11],白色念珠菌中发现了6个,少孢节丛孢菌中发现了13个[7],禾谷炭疽菌中发现了24个[12]。这些已经发现的CFEM蛋白中,白色念珠菌中的Csa2是唯一鉴定了三维(3D)晶体结构的 (图2):该蛋白的CFEM结构域由6个α-螺旋和1个β-链组成,可形成以拉长的N-末端环为柄的螺旋篮状结构[13]。Gong等[12]在禾谷炭疽菌中发现的24个CFEM蛋白中也包含类似于Csa2的螺旋篮状结构,其中6个CFEM蛋白(CgCFEM3、7、8、9、15和20)包含4个α-螺旋,3个CFEM蛋白(CgCFEM4、6和14)包含3个α-螺旋,而CgCFEM1仅包含2个α-螺旋。
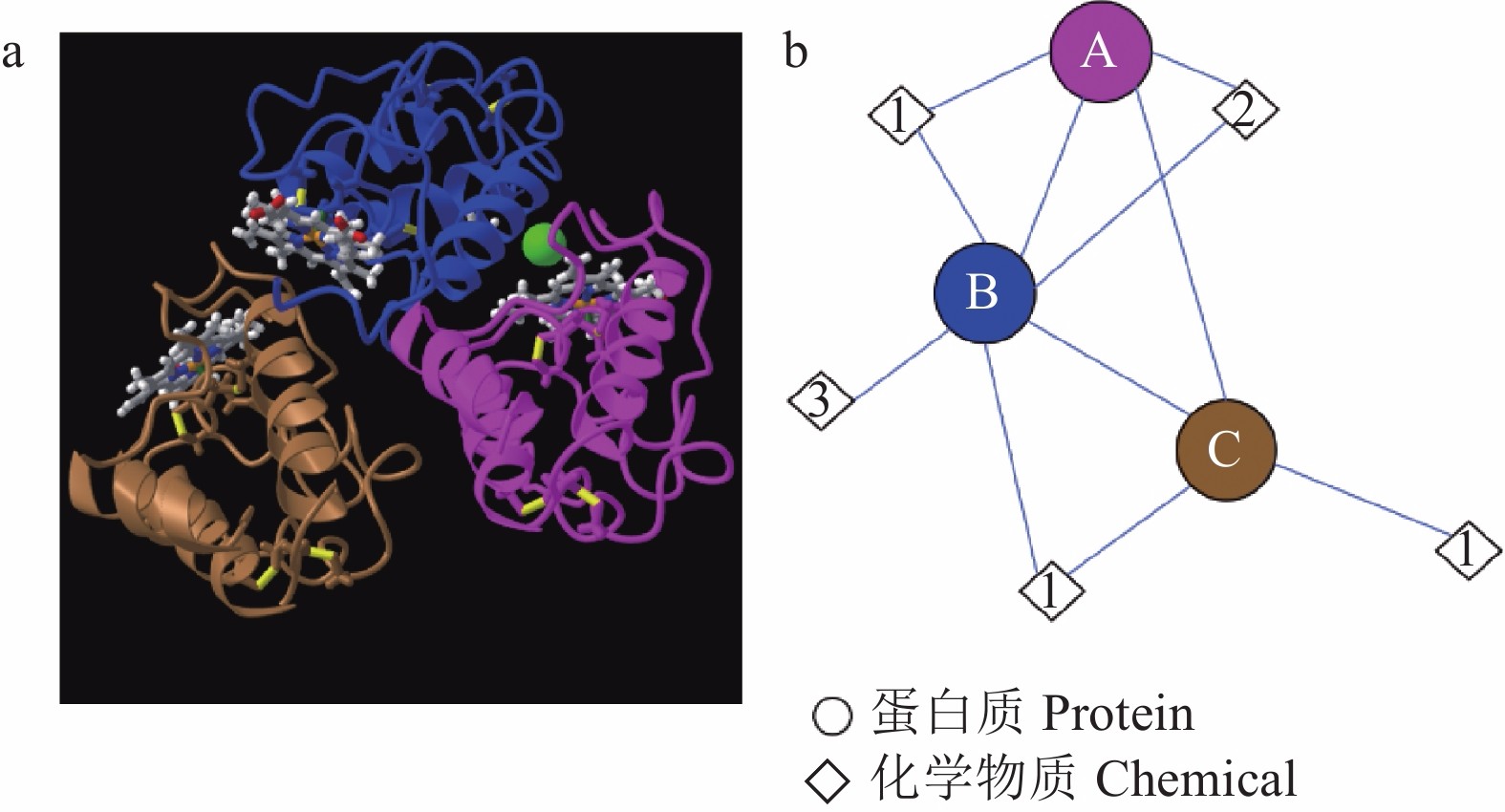
![]() 图 2 白色念珠菌Csa2蛋白的CFEM结构a:Csa2蛋白3个CFEM结构域的3D晶体结构为三聚体(引自NCBI); b:Csa2蛋白的分子组分及它们之间相互作用示意图。A、B、C分别代表3个不同的CFEM结构域,1,2,3分别代表血红素B/C、氯离子和1,2-乙二醇。Figure 2. Structure of CFEM protein domain in Csa2 of Candida albicansa: “3D crystal structure of 3 CFEM protein domains in Csa2 is a trimer” as quoted from NCBI; b: Schematic molecular components and interactions of Csa2. A, B, and C represent 3 CFEM protein domains; 1, 2, and 3 represent heme B/C, chloride ion, and 1,2-ethanediol, respectively.
图 2 白色念珠菌Csa2蛋白的CFEM结构a:Csa2蛋白3个CFEM结构域的3D晶体结构为三聚体(引自NCBI); b:Csa2蛋白的分子组分及它们之间相互作用示意图。A、B、C分别代表3个不同的CFEM结构域,1,2,3分别代表血红素B/C、氯离子和1,2-乙二醇。Figure 2. Structure of CFEM protein domain in Csa2 of Candida albicansa: “3D crystal structure of 3 CFEM protein domains in Csa2 is a trimer” as quoted from NCBI; b: Schematic molecular components and interactions of Csa2. A, B, and C represent 3 CFEM protein domains; 1, 2, and 3 represent heme B/C, chloride ion, and 1,2-ethanediol, respectively.1.2 CFEM蛋白的表达和定位
不同种类真菌中CFEM蛋白的表达部位不同,同一种CFEM蛋白在同一种真菌不同时期的表达量也不同。灰霉病菌(Botrytis cinerea)中的BcCFEM1是一种具有假定GPI锚定位点的CFEM蛋白,大豆接种灰霉病菌后,BcCFEM1的表达水平从0 hpi (hour post-infection)到36 hpi呈上升趋势,之后逐渐下降,但是仍高于0 hpi时的转录水平[14]。在番茄叶片感染灰霉病菌后发现,非G蛋白偶联受体(non-G-protein-coupled receptor,non-GPCR)膜结合CFEM蛋白Bcin07g03260在该菌分生孢子萌发阶段(16 hpi)的表达水平比0 hpi增加了3倍,在坏死生长阶段(48 hpi)表达水平又随之降低[15]。Ling等[16]发现尖孢镰刀菌(Fusarium oxysporum)中属于CFEM家族的基因CFEM_DR1、2、3、4、5和6在侵染的不同时期的表达有所不同。F. oxysporum f. sp. cubense1(Foc1)侵染甘蓝后,CFEM_DR基因的表达分析中可以检测到CFEM_DR2、4、5、6这4个基因的转录水平,但不能检测到CFEM_DR1和CFEM_DR3,可能是因为Foc侵染期间这些基因的表达水平较低。总体而言,这4个CFEM_DR基因的表达在3 dpi(day post-infection)时显著增加。
近平滑假丝酵母中已发现7个CFEM蛋白,其中CFEM2、CFEM3和CFEM6的表达依赖于生物膜和细胞壁调节因子Bcr1,并且这些基因都在低铁条件下被诱导[17]。与这种表达模式相似的还有白色念珠菌中的CFEM蛋白Csa1、Pga7、Pga10和Rbt5[18-19]。Sosinska等研究发现,与pH=4.0的生长条件相比,白色念珠菌在pH=7.0的条件下,Rbt5蛋白的表达水平提高了10倍[20-21]。除此之外,部分CFEM蛋白的表达还与温度有关,例如在可可毛色二孢菌(Lasiodiplodia theobromae)中,与25 ℃和35 ℃时的培养条件相比,LtCFEM1、LtCFEM2、LtCFEM4、LtCFEM5和LtCFEM7在30 ℃时显示出更高的转录水平;与25 ℃相比,在30 ℃和35 ℃下LtCFEM6和LtCFEM8的表达水平均被上调,然而LtCFEM3的表达却被下调[22]。以上结果表明,不同真菌中CFEM蛋白在不同部位、不同时期以及不同生长条件下的表达量不同,预示CFEM蛋白在病原真菌侵染植物时发挥的功能可能是多种多样的。
蛋白质作用的位置与其功能密切相关。为了解CFEM蛋白在细胞中的定位,Gong等[12]选取了禾谷炭疽菌中的10个CFEM效应蛋白在烟草中进行异源表达分析,结果发现大多数CFEM的全长(full length,FL)和去掉信号肽(without signal peptide,NS)部分在细胞中的分布相似。其中CgCFEM1、4、7、9、14、15和20这7种蛋白在细胞膜上被检测到,CgCFEM1 (FL和NS)、CgCFEM4 (NS)和CgCFEM9(FL)在胞质中积累;CgCFEM6和CgCFEM8在细胞核中表达。但也有部分CFEM效应蛋白全长序列与去信号肽序列定位信息不一致,如CgCFEM15 (FL)和CgCFEM20 (FL)定位于细胞膜上,而去掉信号肽之后则在包括细胞膜、细胞核和胞质的整个细胞中表达[12,23]。此外,通过对稻瘟菌中CFEM家族的ACI1蛋白信号肽序列和跨膜区段的分析以及GPI锚定位点的预测,表明它可能是一种膜定位蛋白[24]。该菌中同属于CFEM家族的Pth11和WISH也具有这种特征[25-26]。绿僵菌(Metarhizium anisopliae)中的MaCFEM81、85、87、89和90也属于膜定位蛋白,然而该菌的MaCFEM88在细胞膜和细胞核上均能表达[27]。Wang等[28]发现黄萎病菌(Verticillium dahliae)中的2个CFEM蛋白VdSCP76和VdSCP77,无论是融合红色荧光蛋白还是绿色荧光蛋白,都位于宿主细胞外围,尤其是在质膜上。这表明CFEM蛋白可能在植物体内不同部位发挥作用,明确其在寄主细胞内的定位可以为进一步研究其功能提供可靠的依据。
2. CFEM蛋白的演化
2.1 CFEM蛋白的起源
CFEM结构域至今仅发现存在于真菌蛋白中,在动物、植物和原核生物蛋白中尚未发现该结构[29]。为研究CFEM结构的起源,有研究者对100种真菌的基因组进行CFEM结构搜索,结果显示有64种真菌含有359个CFEM结构的蛋白,主要分布于子囊菌门和担子菌门中,说明CFEM蛋白在真菌中分布较广泛[30-31]。
为了更清楚地了解CFEM结构的分布模式与起源,张真娜[30]构建了担子菌门14个、子囊菌亚门89个和酵母菌亚门10个CFEM蛋白结构域的系统发育关系。结果表明CFEM结构在整个系统发育树中呈分散状态,它们不在同一分类门中聚集出现,这表明CFEM没有独立的起源。也就是说,CFEM结构域的单一起源可以追溯到子囊菌门和担子菌门的最近共同祖先。
2.2 CFEM蛋白的进化
CFEM结构域的形成机制有多种,结构域重复和基因重复是两种最常见的CFEM结构扩增机制,此外还有结构域滑移、基因转换等机制。随着结构域的扩展,其功能也出现了相应的分化[30]。例如,非致病性的酵母菌(Saccharomyces cerevisiae)中只有1个含有CFEM的蛋白CCW14,其参与细胞壁内层的形成[32-33]。致病性真菌尽管往往有多个含CFEM结构的蛋白,但都有一个与CCW14同源的蛋白参与细胞壁的形成,剩下的其他CFEM蛋白则具有不同的功能[7]。这说明CFEM结构最初始的功能只是构成细胞壁或细胞膜的结构组成部分,致病性等功能是在后期的进化过程中产生的。
通过比较不同物种CFEM结构域的长度发现,不同真菌中CFEM的数量存在明显差异[34-35],致病性真菌的CFEM结构蛋白比非致病性真菌的更多。物种自身结构的复杂性以及功能的变化,这些可能是导致CFEM蛋白数目增加的原因。也就是说,越高等的真菌物种含有的CFEM结构域更多,而有些物种所处的环境变化多端,如营寄生生活的真菌或致病性真菌,所含的CFEM结构域数目往往也相应增加,有可能与该结构域维持物种基本的生存能力有关系[7]。
3. CFEM蛋白的功能及作用机制
3.1 维持真菌胞内所需铁含量
几乎所有植物、细菌和真菌生长发育都需要铁营养元素[36]。在病原微生物侵染寄主植物时,病原微生物及其寄主都需要铁离子来保持各自的生理功能、生长和增殖。因此,铁离子信号影响植物与病原微生物之间的互作关系。目前已在多个真菌体内发现了铁吸收系统,例如,铁载体调控的Fe3+吸收、还原铁同化作用、亚铁血红素吸收或者Fe2+直接吸收[37]。
一些研究表明,CFEM蛋白在维持真菌细胞内铁含量方面发挥重要的作用。例如,白色念珠菌中的高度同源的两个蛋白Rbt5和Rbt51是CFEM家族的一种细胞外GPI锚定血红素结合蛋白,可促进血红素铁的摄取。Rbt5是白色念珠菌利用血红素和血红蛋白作为铁源的关键因子,缺失会引起严重的缺铁表型[17]。Pga7的缺失会引起比Rbt5缺失更严重的缺铁表型,表明这两个蛋白质都能有效地帮助白色念珠菌从血红蛋白中获取铁元素[38]。其中Rbt5对白色念珠菌的致病性并没有明显作用,而Pga7是该菌致病所必需的[38-39]。酵母菌中虽然缺乏血红蛋白受体Rbt5的直系同源物,但是在其体内异源表达白色念珠菌中的Rbt51时,同样也能使其将血红蛋白作为铁源[10]。除此之外,Csa2是白色念珠菌中Rbt5蛋白家族和CFEM蛋白家族的成员,有研究证明Csa2在白色念珠菌菌丝生长过程中参与利用血红蛋白中的铁元素[40]。Csa2蛋白的第80位天冬氨酸残基能帮助C. albicans获取血红素中的Fe3+,如果该突变位点为组氨酸残基,则该菌获取的是血红素中的Fe2+[13]。Srivastava等[41]首次证明了光滑假丝酵母(Candida glabrata)中属于CFEM家族的细胞壁结构蛋白CgCcw14和假定溶血素CgMam3,对维持细胞内铁含量至关重要。
3.2 调控真菌的生长发育
Ling等[16]研究发现尖孢镰刀菌的CFEM_DR蛋白在菌丝体中的表达比在分生孢子中的表达要高,并且一些CFEM_DR基因在菌丝体生长晚期的表达比早期更高,表明CFEM_DR基因在尖孢镰刀菌菌丝体生长中具有重要作用。而且尖孢镰刀菌能穿透寄主根的皮层组织进入维管组织,并通过菌丝体的生长与液流一起运输。灰霉病菌中的BcCFEM1敲除后,虽然∆Bccfem1突变体分生孢子的萌发没有受到影响,但其分生孢子产量明显减少[14]。CFEM蛋白Bcin07g03260的缺失,导致∆CFEM-Bcin07g03260突变体分生孢子萌发率明显降低,同时芽管长度显著缩短[15]。
稻瘟菌中被命名为WISH的蛋白同时具有CFEM和GPCR结构。该基因调控稻瘟菌的菌丝生长和分生孢子分化与形成,此外,还在维持细胞壁完整性、对宿主的表面感应和真菌表面疏水性方面发挥调控作用。敲除WISH后,∆wish突变体分生孢子产量明显增加并且无法形成附着胞进而导致无法侵染寄主,这表明WISH蛋白负调控稻瘟菌分生孢子的产生且是该菌附着胞形成所必需的[42]。
希金斯炭疽菌(Colletotrichum higginsianum Sacc.)中含有CFEM结构域的效应因子ChEP113被敲除后,∆ChEP113突变体菌株气生菌丝增多,菌丝生长变慢且形态明显改变:末端笔直、分叉减少,孢子产量和菌丝重量明显增加;孢子萌发时大多只在一侧产生芽管,且只有少数分生孢子可以正常萌发并产生黑化附着胞,这表明ChEP113蛋白参与调控希金斯炭疽菌的生长发育,在该菌菌落形态、菌丝生长、产孢和孢子萌发方面发挥重要作用[43]。
3.3 影响真菌致病力
已有研究表明,真菌在侵染阶段表达的分泌蛋白更有可能影响病原体的致病性[44]。Wang等通过研究表明在侵染阶段玉米大斑病菌(Setosphaeria turcica)中包含StCFEM3、4、5,10、12和13这6个效应蛋白在内的13个CFEM蛋白都有表达,这表明玉米大斑病菌中的StCFEMs可能起着致病因子的作用[45]。尖孢镰刀菌中CFEM_DR基因在侵染寄主后第3天在寄主根中大量表达,而该蛋白已经被证实了在宿主的定植和侵染中发挥重要的致病作用[16]。小新壳梭孢菌(Neofusicoccum parvum)中含有CFEM结构域的蛋白NpCysRP5也是在早期侵染阶段高度表达[46]。此外,还有灰霉病菌中的BcCFEM1[14]、尖孢镰刀菌中的CFEM_DR基因[16]、C. graminicola中22个含有CFEM结构域的效应因子[12]以及可可毛色二孢菌中除LtCFEM2和LtCFEM6外的LtCFEMs等[22],并且在烟草中异源表达BcCFEM1、LtCFEM1会导致叶片明显萎黄[14,22]。Chen等[47]通过研究发现禾谷镰刀菌(Fusarium graminearum)中的FgCFEM11和FgCFEM23有可能在侵染植物期间作为效应因子参与致病过程。小麦叶锈菌(Puccinia triticina)中的CFEM效应因子PTTG-08198加速了由BAX触发的植物细胞死亡并且诱导了烟草细胞中活性氧(Reactive Oxygen Species,ROS)的积累[48]。MoCDIP2和MoCDIP11是稻瘟菌中诱导寄主细胞死亡和引发水稻防御反应的CFEM效应因子[49-50],而禾谷炭疽菌中具有CFEM结构域的GLRG_01006.1却是能抑制细胞死亡的效应因子[23]。
为进一步研究CFEM蛋白对真菌毒力的影响,Zhu等对BcCFEM1进行了敲除,结果表明突变体菌株表现出对寄主的致病性降低以及对过氧化氢的敏感性增强。推测突变体的致病性降低可能是其对植物防御相关活性氧高度敏感性的结果[14]。核盘菌(Sclerotinia sclerotiorum)中的SsCFEM1被敲除后,突变体菌株毒力也明显下降[51]。Peng等[52]通过局部感染和注射蜡蛾幼虫来评估白僵菌(Beauveria bassiana)的毒力,结果显示野生型菌株杀死了所有幼虫,而∆Bbcfem7和∆Bbcfem8突变体菌株对蜡蛾幼虫的致死率不到50%,表明BbCFEM7和BbCFEM8显著促进了该菌的毒力。以上结果都说明了CFEM蛋白对真菌病原物的重要性,同时也表明CFEM蛋白可能作为毒性致病因子参与调控真菌致病性相关的重要生理过程。但不是所有的CFEM蛋白都和真菌致病性有关,例如CfmA-C可能与烟曲霉细胞壁的稳定性有关,但其缺失不影响该菌的致病力[8],还有CCW14是在非致病性真菌中发现的[32-33]。
3.4 促进真菌寄生
在白色念珠菌中,菌丝体的生长和生物膜的形成能力被认为是重要的毒力特征,Pérez等的研究中表明,Csa1、Pga7和Rbt5在菌丝中上调表达,并且编码这类蛋白的基因被敲除后,突变体在生物膜形成过程中存在缺陷[53]。这表明这些蛋白可能作为信号分子和毒性致病因子在病原菌侵染过程中发挥作用。除此之外,在稻瘟菌中,WISH被敲除后,附着胞发育明显被抑制,无法识别寄主表面和穿透水稻叶片,导致其致病性几乎丧失,这表明WISH蛋白参与宿主表面识别和真菌致病性[43]。ACI1为胞外受体信号分子,通过与腺苷酸环化酶(MAC1)互作,参与环磷酸腺苷(cyclic adenosine monophosphate,cAMP)信号传递,直接影响稻瘟菌附着胞的形成[54-55]。同样地,Pth11作为稻瘟菌附着胞及芽管胞内信号分子,被敲除后,突变体表现出附着胞形成延迟以及致病性减弱[11,56]。CFEM作为一种膜蛋白,可以充当细胞表面受体或信号转导因子,或者在宿主-病原体互作中充当粘附分子[6],因此,推测CFEM结构域蛋白可能通过这些途径发挥作用从而促进真菌寄生。
4. 问题与展望
CFEM作为真菌特有的蛋白结构,其在不同真菌中的分布数有较大差异,说明CFEM蛋白生物学功能存在较大进化与差异。CFEM蛋白除了最初的功能参与细胞壁形成有关外,目前报道的CFEM蛋白功能主要体现在维持真菌胞内所需铁含量,参与真菌生长发育过程,促进病原真菌致病及抑制寄主免疫反应等。然而,CFEM的研究存在以下几方面问题有待进一步解决:尽管CFEM蛋白在真菌中分布广泛,然而关于CFEM蛋白功能的研究目前大多数只限于稻瘟菌、禾谷炭疽菌、白色念珠菌等少数真菌,在其他真菌中的研究非常的少;其次,CFEM蛋白的功能逐渐被人们揭示,然而关于其作用机理、作用靶标的研究还存在较大空白,需进一步增加有关CFEM蛋白功能研究的深度,解析CFEM蛋白的功能分子机制;此外,虽然CFEM结构域的长度与氨基酸序列具有高度的保守性,但不同真菌物种CFEM结构域的数量存在显著差异,其功能的多样性是否与真菌体内CFEM蛋白数量以及结构域进化有关。这些问题都需要进一步的深入研究来阐明。通过揭示CFEM蛋白的功能,其作用靶标与上下游信号网络及其起源和分化,将有助于阐明CFEM蛋白的生理功能,进一步丰富植物-病原物相互作用机理,为作物病害防控策略的制定提供参考。
-
![]()
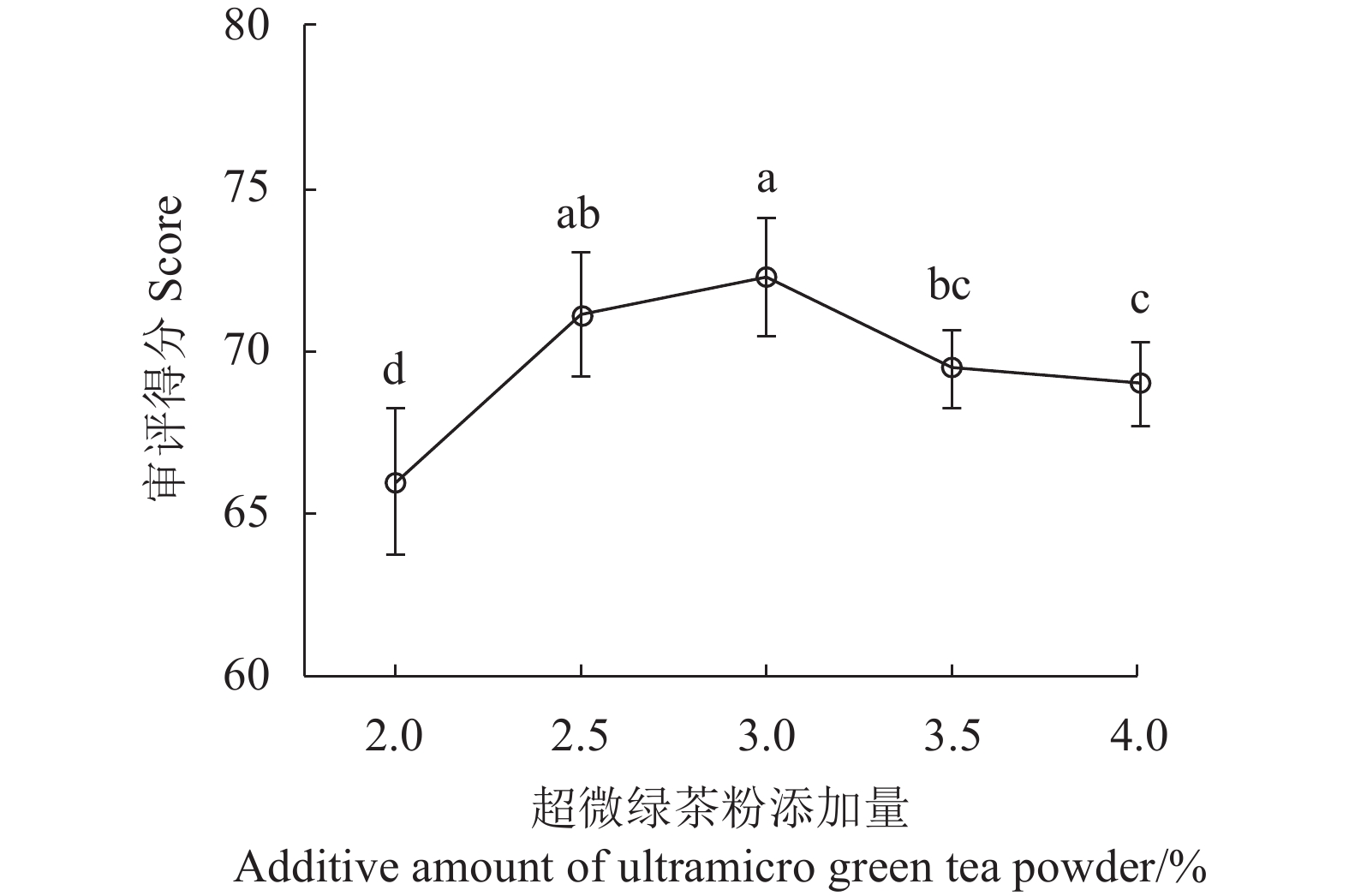
图 1 超微绿茶粉添加量对曲奇的影响
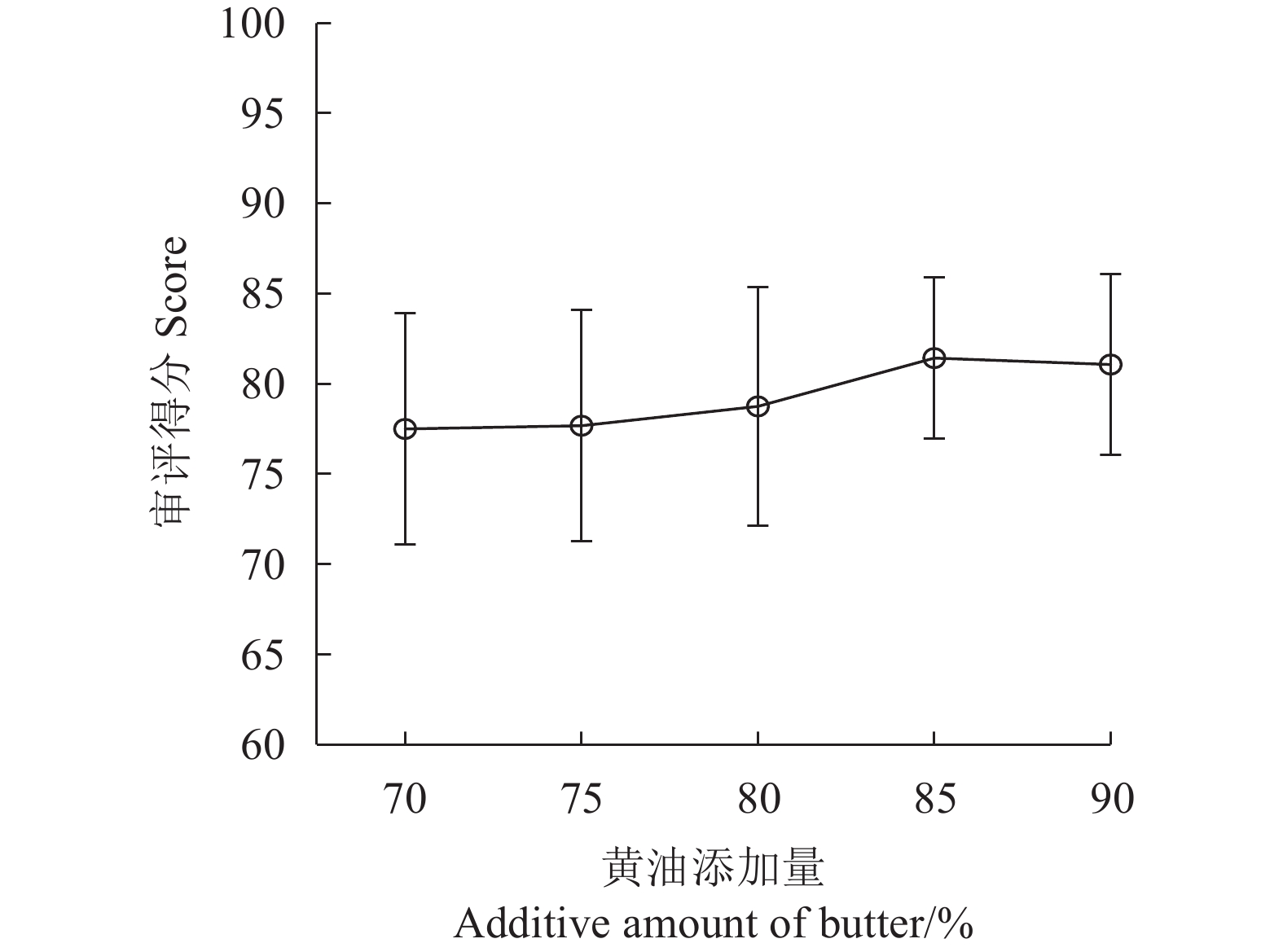
图中不同小写字母表示显著差异(P<0.05),相同或无小写字母表示差异不显著(P>0.05)。图2、3同。
Figure 1. Effect of quantity of added ultramicro green tea powder on cookie quality
Data with different lowercase letters indicate significant differences (P<0.05); those with same or no lowercase letters, no significant difference (P>0.05). Same fo Figs.2,3.
表 1 试验变量的因素和水平
Table 1 Factors and levels of test variables
水平
Level因素 Factor 超微绿茶粉添加量A
Amount of ultramicro green tea powder/%黄油添加量B
Amount of butter/%糖粉添加量C
Amount of powdered sugar/%−1 2.5 80 24 0 3.0 85 26 1 3.5 90 28  下载: 导出CSV
下载: 导出CSV
表 2 超微绿茶粉曲奇饼干审评标准
Table 2 Evaluation criteria of ultramicro green tea powder cookie
项目
Item评分标准
Standard for evaluation得分
Score形态
Form外形完整、饱满,不收缩、不变形、不起泡,厚薄均一,大小均匀,底部平整,无粘连,边缘平滑 16~20 外形较完整、较饱满、收缩和变形少、起泡少,厚薄较均一,大小较均匀,底部平整,无粘连,边缘较平滑 10~15 外形不完整、有破碎、凹底和收缩较多、表面起泡,厚薄不均一,大小不均匀,有粘连 <10 色泽
Color表面呈草绿色,有光泽,无色泽不均,无白粉 16~20 表面呈草绿色,有光泽,色泽基本均匀,边缘和底部略有棕黄色,但不出现焦黑或过白 10~15 光泽不足、不均匀,有焦黑色或过白现象 <10 组织
Texture断面结构细密均匀,无杂质,无较大孔洞,表面细腻 16~20 断面结构略粗糙、不均匀,无杂质,有少量孔洞,表面较细腻 10~15 断面结构粗糙、不均匀,有杂质,有较多孔洞,表面亦粗糙 <10 滋味
Flavor口感酥松、细腻、不粘牙,甜而不腻 16~20 口感较酥松、较细腻、略粘牙,甜而不腻 10~15 口感不酥松、粗糙、粘牙,口味过甜或淡无味 <10 风味
Relish茶香味浓郁,与饼干香味协调,无苦涩味, 16~20 有茶香味,略有苦涩味 10~15 无明显茶香,苦涩味较重 <10
下载: 导出CSV
表 3 响应面分析试验结果
Table 3 Design and results of response surface analysis
试验号
Serial numberA超微绿茶粉
添加量
Amount of
ultramicro green
tea powderB黄油
添加量
Amount of
butterC糖粉
添加量
Amount of
powdered
sugarY感官审
评得分
Sensory scores1 0(3%) 0(85%) 0(26%) 85.83 2 −1(2.5%) 0 1(28%) 73.33 3 0 0 0 86.67 4 0 −1(80%) −1(24%) 78.23 5 −1 1(90%) 0 72.50 6 1(3.5%) 0 1 70.67 7 0 0 0 84.67 8 −1 −1 0 75.67 9 0 −1 1 76.13 10 0 1 1 73.03 11 0 0 0 85.83 12 1 −1 0 71.26 13 0 1 −1 76.03 14 1 1 0 70.67 15 1 0 −1 71.17 16 −1 0 −1 73.13 17 0 0 0 87.00
下载: 导出CSV
表 4 回归方程的方差分析
Table 4 Analysis of variance of regression equation
方差来源
Source of variance平方和
Sum of square自由度
Degree of freedom均方
Mean squareF值
F valueP值
P value模型
Model617.54 9 68.62 70.84 <0.0001 A 超微绿茶粉
Ultramicro green tea powder14.74 1 14.74 15.22 0.0059 B 黄油
Butter10.26 1 10.26 10.59 0.0140 C 糖粉
Powdered sugar3.65 1 3.65 3.76 0.0935 AB 1.66 1 1.66 1.72 0.2313 AC 0.12 1 0.12 0.13 0.7326 BC 0.20 1 0.20 0.21 0.6614 A2 313.41 1 313.41 323.57 <0.0001 B2 98.94 1 98.94 102.15 <0.0001 C2 118.16 1 118.16 121.99 <0.0001 残差项
Residual6.78 7 0.97 失拟项
Lack of fit3.50 3 1.17 1.43 0.3591 纯误差项
Pure error3.28 4 0.82 总误差项
Cor total624.32 16 相关系数R2 0.9897 校正决定系数Radj2 0.9752
下载: 导出CSV
表 5 超微绿茶粉曲奇饼干与普通曲奇饼干的质构参数
Table 5 Texture measurements on ultramicro green tea powder cookie and common cookies
质构参数
Parameter of texture analyzer超微绿茶粉曲奇饼干
Ultramicro green tea powder cookies普通曲奇饼干
Cookies硬度
Rigidity/g578.44±37.93* 467.59±66.92 酥性
Short/(g·s)626.46±67.58 509.60±97.56 *表示同行数据之间差异显著(P<0.05)。
* indicates that values in the same row are with significant differences (P<0.05).
下载: 导出CSV
-
[1] 刘仲华, 陈宗懋, 杨亚军, 等. 创新驱动中国茶产业高质量发展: 从茶学基础研究到支撑产业发展 [J]. 中国茶叶, 2021, 43(2):1−9. LIU Z H, CHEN Z M, YANG Y J, et al. Innovation drives high-quality development of Chinese tea industry—From basic research of tea science to supporting industrial development [J]. China Tea, 2021, 43(2): 1−9.(in Chinese)
[2] 宛晓春. 茶叶生物化学[M]. 3版. 北京: 中国农业出版社, 2003. [3] 任艳军, 杜彬, 宋晓飞. 不同加工条件对茶叶超微粉碎效果的影响 [J]. 河北科技师范学院学报, 2009, 23(2):64−66. DOI: 10.3969/j.issn.1672-7983.2009.02.015 REN Y J, DU B, SONG X F. Effect of different processing conditions on tea ultrafine pulverizing [J]. Journal of Hebei Normal University of Science & Technology, 2009, 23(2): 64−66.(in Chinese) DOI: 10.3969/j.issn.1672-7983.2009.02.015
[4] 李琳, 刘天一, 李小雨, 等. 超微茶粉的制备与性能 [J]. 食品研究与开发, 2011, 32(1):53−56. DOI: 10.3969/j.issn.1005-6521.2011.01.016 LI L, LIU T Y, LI X Y, et al. Processing and properties of ultrafine tea powder [J]. Food Research and Development, 2011, 32(1): 53−56.(in Chinese) DOI: 10.3969/j.issn.1005-6521.2011.01.016
[5] 隋秀芳, 赵鹏, 王玉珠, 等. 超微绿茶粉营养功能成分分析及其应用 [J]. 食品研究与开发, 2012, 33(10):173−177. SUI X F, ZHAO P, WANG Y Z, et al. Research on nutrition and function components and application of fine green tea powder [J]. Food Research and Development, 2012, 33(10): 173−177.(in Chinese)
[6] 李林秀. 超微茶粉及茶叶糕点加工技术 [J]. 现代园艺, 2016(2):222−223. DOI: 10.3969/j.issn.1006-4958.2016.02.173 LI L X. Processing technology of ultrafine tea powder and tea cake [J]. Xiandai Horticulture, 2016(2): 222−223.(in Chinese) DOI: 10.3969/j.issn.1006-4958.2016.02.173
[7] 于克学, 孙建霞, 白卫滨, 等. 超微茶粉面条的研制 [J]. 食品科技, 2008, 33(6):121−123. DOI: 10.3969/j.issn.1005-9989.2008.06.037 YU K X, SUN J X, BAI W B, et al. Study on ultramicrosome green tea powder noodle [J]. Food Science and Technology, 2008, 33(6): 121−123.(in Chinese) DOI: 10.3969/j.issn.1005-9989.2008.06.037
[8] 祁国栋, 张炳文. 超微粉碎技术在中低档茶叶食品开发中的应用 [J]. 农业工程技术(农产品加工业), 2008(9):33−35. QI G D, ZHANG B W. Application of superfine grinding technology in the development of middle and low-grade tea food [J]. Agriculture Engineering Technology (Agricultural Product Processing Industry), 2008(9): 33−35.(in Chinese)
[9] 金寿珍. 超微茶粉加工技术 [J]. 中国茶叶, 2007, 29(6):12−14. DOI: 10.3969/j.issn.1000-3150.2007.06.005 JIN S Z. Processing technology of superfine tea powder [J]. China Tea, 2007, 29(6): 12−14.(in Chinese) DOI: 10.3969/j.issn.1000-3150.2007.06.005
[10] 国家质量监督检验检疫总局, 中国国家标准化管理委员会. 饼干: GB/T 20980—2007[S]. 北京: 中国标准出版社, 2008. [11] 李博, 刘明理, 陈伯玮, 等. 绿茶曲奇饼干的研制及配方优化 [J]. 农产品加工(学刊), 2013(8):42−44. LI B, LIU M L, CHEN B W, et al. Preparation of green tea cookies and the optimization of its formula [J]. Academic Periodical of Farm Products Processing, 2013(8): 42−44.(in Chinese)
[12] 温晓菊, 张维, 王英, 等. 茶曲奇的研制 [J]. 热带农业科学, 2013, 33(9):55−59. DOI: 10.3969/j.issn.1009-2196.2013.09.013 WEN X J, ZHANG W, WANG Y, et al. Development of cookie made with tea [J]. Chinese Journal of Tropical Agriculture, 2013, 33(9): 55−59.(in Chinese) DOI: 10.3969/j.issn.1009-2196.2013.09.013
[13] 张新富, 王玉, 杨绍兰, 等. 绿茶曲奇饼干的研制 [J]. 食品工业科技, 2009, 30(5):278−279,282. DOI: 10.13386/j.issn1002-0306.2009.05.063 ZHANG X F, WANG Y, YANG S L, et al. Study on development of cookie made with green tea [J]. Science and Technology of Food Industry, 2009, 30(5): 278−279,282.(in Chinese) DOI: 10.13386/j.issn1002-0306.2009.05.063
[14] 雷文平, 吴诗敏, 李彩虹, 等. 响应面法优化凝固型发酵椰奶工艺 [J]. 中国酿造, 2019(2):212−216. DOI: 10.11882/j.issn.0254-5071.2019.02.041 LEI W P, WU S M, LI C H, et al. Process optimization of set-style fermented coconut milk by response surface methodology [J]. China Brewing, 2019(2): 212−216.(in Chinese) DOI: 10.11882/j.issn.0254-5071.2019.02.041
[15] 张翊, 黄岚. 响应面法优化枸杞富硒曲奇饼干配方 [J]. 粮食与油脂, 2018, 31(8):50−54. DOI: 10.3969/j.issn.1008-9578.2018.08.014 ZHANG Y, HUANG L. Optimization of Lycium barbarum selenium-rich cookies by response surface methodology [J]. Cereals & Oils, 2018, 31(8): 50−54.(in Chinese) DOI: 10.3969/j.issn.1008-9578.2018.08.014
[16] 张强, 赵卉珉, 梁进. 响应面法优化戊聚糖曲奇饼干工艺配方研究 [J]. 粮油食品科技, 2019, 27(1):24−29. DOI: 10.16210/j.cnki.1007-7561.2019.01.005 ZHANG Q, ZHAO H M, LIANG J. Study on optimization of the technological formulation of pentosan cookie by response surface method [J]. Science and Technology of Cereals, Oils and Foods, 2019, 27(1): 24−29.(in Chinese) DOI: 10.16210/j.cnki.1007-7561.2019.01.005
[17] 王慧玲, 刘雪君, 丁芳, 等. 响应面法优化绿茶调味酱配方 [J]. 包装与食品机械, 2017, 35(4):30−36. DOI: 10.3969/j.issn.1005-1295.2017.04.007 WANG H L, LIU X J, DING F, et al. Optimization of green tea flavor paste by response surface methodology [J]. Packaging and Food Machinery, 2017, 35(4): 30−36.(in Chinese) DOI: 10.3969/j.issn.1005-1295.2017.04.007
[18] 徐易, 邓尚贵, 霍健聪. 响应面法在微胶囊鱼油曲奇饼干研制中的应用 [J]. 食品工业, 2015, 36(9):160−164. XU Y, DENG S G, HUO J C. Application of response surface methodology in the development of fish oil microcapsules cookie [J]. The Food Industry, 2015, 36(9): 160−164.(in Chinese)
[19] 胡玥明, 孙桂菊, 姚宏亮. 响应面法优化鸡小肠黏膜唾液酸提取工艺 [J]. 福建农业学报, 2020, 35(6):665−672. DOI: 10.19303/j.issn.1008-0384.2020.06.013 HU Y M, SUN G J, YAO H L. Response surface optimization of sialic acid extraction from chicken small-intestine mucosa [J]. Fujian Journal of Agricultural Sciences, 2020, 35(6): 665−672.(in Chinese) DOI: 10.19303/j.issn.1008-0384.2020.06.013
[20] 阮怿航, 吴亮宇, 鲁静, 等. 低共熔溶液提取铁观音茶多酚工艺的响应面法优化 [J]. 福建农业学报, 2020, 35(2):217−225. DOI: 10.19303/j.issn.1008-0384.2020.02.013 RUAN Y H, WU L Y, LU J, et al. Deep eutectic solvents extraction of polyphenols from tieguanyin tea optimized by response surface method [J]. Fujian Journal of Agricultural Sciences, 2020, 35(2): 217−225.(in Chinese) DOI: 10.19303/j.issn.1008-0384.2020.02.013
[21] 蒋旖旋, 龚超, 侯莉莉, 等. 响应面-主成分分析法研制莲房膳食纤维曲奇 [J]. 食品研究与开发, 2016, 37(1):100−106. DOI: 10.3969/j.issn.1005-6521.2016.01.025 JIANG Y X, GONG C, HOU L L, et al. Development of lotus seed pot dietary fiber cookie by response surface methodology and principal component analysis [J]. Food Research and Development, 2016, 37(1): 100−106.(in Chinese) DOI: 10.3969/j.issn.1005-6521.2016.01.025
[22] AHMAD M, BABA W N, WANI T A, et al. Effect of green tea powder on thermal, rheological & functional properties of wheat flour and physical, nutraceutical & sensory analysis of cookies [J]. Journal of Food Science and Technology, 2015, 52(9): 5799−5807. DOI: 10.1007/s13197-014-1701-3
[23] 中国茶叶流通协会. 2021年中国茶叶产销形势报告 [J]. 茶世界, 2022(3):1−18. China Tea Marketing Association. Situation report of Chinese tea production and marketing in 2021 [J]. Tea World, 2022(3): 1−18.(in Chinese)
[24] 陈宗懋. 科技创新和我国茶产业发展[C]//中国茶叶学会年会论文集, 2015: 6-13. CHEN Z M. Innovation of scientific and technological and the development of chinese tea industry[C]//China Tea Science Society, 2015: 6-13. (in Chinese)
[25] 管曦, 杨江帆, 谢向英, 等. 中国茶产业供求失衡的再思考: 基于国内主要茶叶市场数据的分析 [J]. 茶叶学报, 2017, 58(2):75−79. DOI: 10.3969/j.issn.1007-4872.2017.02.008 GUAN X, YANG J F, XIE X Y, et al. Over-supply of tea in China—a marketing analysis [J]. Acta Tea Sinica, 2017, 58(2): 75−79.(in Chinese) DOI: 10.3969/j.issn.1007-4872.2017.02.008
[26] 董海洲, 刘传富, 侯汉学. 绿茶月饼加工工艺参数及其理化特性 [J]. 食品与发酵工业, 2003, 29(8):88−91. DOI: 10.3321/j.issn:0253-990X.2003.08.020 DONG H Z, LIU C F, HOU H X. Processing parameters and physicochemical properties of green tea moon cakes [J]. Food and Fermentation Industries, 2003, 29(8): 88−91.(in Chinese) DOI: 10.3321/j.issn:0253-990X.2003.08.020
[27] 叶丹榕, 彭小燕, 蔡香珍, 等. 响应面法优化茉莉花茶风味茶油桃酥配方 [J]. 热带农业科学, 2020, 40(7):95−102. YE D R, PENG X Y, CAI X Z, et al. Optimization of formula of jasmine tea flavor tea-seed oil crisp cakes based on response surface methodology [J]. Chinese Journal of Tropical Agriculture, 2020, 40(7): 95−102.(in Chinese)
[28] 孙科祥, 计红芳, 张令文, 等. 茶香绿豆糕的研制 [J]. 食品研究与开发, 2009, 30(8):44−47. DOI: 10.3969/j.issn.1005-6521.2009.08.013 SUN K X, JI H F, ZHANG L W, et al. Processing technique for mung bean cake with biluochun tea [J]. Food Research and Development, 2009, 30(8): 44−47.(in Chinese) DOI: 10.3969/j.issn.1005-6521.2009.08.013
-
期刊类型引用(1)
1. 王艳玲,郭勇,高洁. 病原真菌CFEM蛋白功能及分子机制研究进展. 华中农业大学学报. 2025(01): 156-167 .  百度学术
百度学术
其他类型引用(2)


计量
- 文章访问数: 504
- HTML全文浏览量: 166
- PDF下载量: 69
- 被引次数: 3
