

Functions of Endogenous Hormones in Regulating Bud Dormancy of Pear Flowers
-
摘要:目的 分析砂梨花芽休眠进程中内源激素的含量及平衡关系对梨花芽休眠进程的调控作用,为研究南方短低温梨花芽休眠解除的调控机理奠定基础。方法 以黄花梨和蜜雪梨2个砂梨品种花芽为试材,利用HPLC技术分析赤霉素(GA3)、脱落酸(ABA)、3-吲哚乙酸(IAA)、玉米素(ZT)等4种内源激素含量以及GA3/ABA、IAA/ABA、ZT/ABA、(IAA+GA3)/ABA比值在2个砂梨品种花芽休眠进程中的变化。结果 在砂梨花芽休眠进程中,GA3和ABA的影响主要在内休眠解除启动期,并且GA3促进了花芽内休眠解除启动,而ABA抑制了花芽休眠解除启动;IAA在休眠进程中的变化规律不明显,对于休眠解除的影响不显著;ZT在黄花梨中促进了内休眠的解除,而在蜜雪梨中则作用于内休眠解除的启动。GA3/ABA和(IAA+GA3)/ABA在砂梨花芽休眠进程中变化趋势基本相同,均表现出在内休眠阶段比值较小,并且变化趋势不明显,而内休眠解除启动时明显升高,并且在内休眠解除时稍低于内休眠解除启动期;IAA/ABA其对休眠进程的影响较小;ZT/ABA的比值在黄花梨花芽休眠解除启动期稍低于内休眠解除期,而在蜜雪梨中花芽休眠解除启动期会稍高于内休眠解除期。结论 砂梨花芽休眠进程中GA3和ABA两种內源激素起主要作用,ZT不能直接影响休眠进程,而且内源激素调控花芽的休眠进程更主要取决于各内源激素的平衡。不同品种之间內源激素变化不完全一致,但总的趋势相似,本研究为內源激素调控休眠的研究奠定了基础。Abstract:Objective Functions of endogenous hormones in Pyrus pyrifolia on regulating the flower bud dormancy were studied.Method Flower buds of P. pyrifolia Huanghua and P. pyrifolia Mixue in various stages of dormancy were collected to determine the contents of GA3, ABA, IAA, and ZT as well as the ratios of GA3/ABA, IAA/ABA, ZT/ABA, and (IAA+GA3)/ABA by HPLC.Result GA3 and ABA exerted significant effects on the bud dormancy mostly during the endodormancy-releasing/bud-germinating stage as GA3 initiated but ABA inhibited the release. IAA showed no significant effect in that regard, and ZT in Huanghua promoted the release, while ZT in Mixue initiated the dormancy. The ratios of GA3/ABA and (IAA+GA3)/ABA of both pear varieties were similarly low during dormancy. They rose significantly when the dormancy-release started and slightly declined afterward. The IAA/ABA ratio changed little, whereas the ZT/ABA decreased slightly in Huanghua, but the opposite in Mixue, at the initiation of dormancy-release.Conclusion GA3 and ABA played a major role in the dormancy of the pear flower buds, but ZT did not directly affect the dormancy-release. The process appeared to be regulated by an integrated effect of balance among these endogenous hormones, rather than individually. Although the changes on the endogenous hormones during flower bud dormancy differed between the two pear varieties, a similar general trend seemed existed.
-
Keywords:
- Pyrus pyrifolia /
- endogenous hormones /
- GA3 /
- ABA /
- dormancy
-
0. 引言
【研究意义】梨(Pyrus L.)是蔷薇科(Rosaceae)落叶果树,是世界上的主要栽培果树之一。随着全球气候变暖,一些地区由于需冷量的不足导致梨树不能及时打破休眠,给梨树的后续生长及产量带来了不良影响[1]。目前在中国南方砂梨品种群普遍存在“二次花”现象,而作为短低温需求的蜜雪梨二次花发生频率在70%左右,严重影响了砂梨在南方地区的推广种植[2],所以在中国南方地区研究梨休眠对推动梨产业的发展具有重要意义。【前人研究进展】植物激素对于植物休眠进程具有调控作用,并且不同激素的作用不同,赤霉素(GA3)和脱落酸(ABA)被认为是影响植物芽休眠最重要的激素[3]。普遍认为GA3是休眠解除物质,能够代替低温促进内休眠的解除[4,5],但是刘国琴[3]指出,GA3与果树萌芽关系十分密切,认为GA3不是休眠解除的必需因素,而是芽萌发的必要条件。ABA被认为是休眠激素,是促进休眠和抑制萌发的激素[6]。多数研究结果表明,ABA 含量降低促进落叶果树芽在适宜环境条件下的萌芽[7]。3-吲哚乙酸(IAA)一般被认为不直接参与植物芽休眠的调控[6],郑国生等[8]通过对牡丹芽休眠的研究,揭示了IAA并不是芽休眠解除的必要因子,而是芽萌动的必要因子。IAA在休眠解除期间含量会增加,但并不直接参与休眠调控[4,6,9,10],并且许多研究指出低浓度的IAA能够促进芽的萌发,这可能是因为低浓度的IAA能够促进根系的生长,从而促进根系中细胞分裂素的合成,促进了萌芽物质向地上运输,间接促进了休眠解除,而高浓度的IAA抑制芽的萌发主要是因为顶端优势的作用[9]。细胞分裂素(CTK)主要是以细胞分裂素偶联物的形式通过木质部在植物体内运输,除了可以刺激细胞的分裂分化以外,还具有打破休眠的作用[9],相关研究证明CTK具有打破植物休眠的效果[8,11,12]。而玉米素(ZT)是细胞分裂素的一种。袁燕波等[12]研究表明GA3/ABA 和ZT/ABA 的比值随休眠的加深而减小,随休眠的解除而增大。高庆玉等[13]在树莓上的研究表明IAA/ABA的变化趋势与GA3/ABA变化趋势相同,说明ABA调控休眠解除与其他激素的动态平衡有关。段成国等[14]在研究甜樱桃时发现ABA/GA3、ABA/ZT的比值随休眠进程的深入而升高,随着休眠解除而下降。这些研究均说明了內源激素的平衡对植物芽休眠解除的重要性[14]。【本研究切入点】休眠的起始、终止以及休眠各阶段的变化均受激素调节。内源激素在落叶果树上的研究主要集中在桃[5]、甜樱桃[14],苹果[15]、葡萄[6]等,而与梨休眠相关的激素研究还较少[16],并且内源激素作为植物体内庞大的调控网络,其对落叶果树休眠解除的调控机理尚未阐明清楚。【拟解决的关键问题】本研究以黄花梨和蜜雪梨侧花芽为试验材料,利用HPLC技术分析梨花芽休眠进程中内源激素的含量及平衡关系,试图探讨内源激素对于梨花芽休眠进程的调控机理,为解决南方短低温梨花芽休眠解除的调控机理奠定基础。
1. 材料与方法
1.1 试验材料
本试验供试品种为黄花梨(Pyrus pyrifolia. Huanghua) 和蜜雪梨(Pyrus pyrifolia.Mixue),黄花梨花芽采自福建省建宁县,蜜雪梨花芽采自福建省上杭县,采样用树均为10年左右的成年结果树,在采样过程中试验用树不进行任何化学处理和修剪。每次采样设置3次生物学重复(每5株为1次生物学重复),采集花芽用湿布包好,带回实验室,选取饱满的侧花芽除去外部鳞片和绒毛,于−80 ℃冰箱保存备用。
每个年度采样时间及休眠进程如表1所示,休眠进程判定方法参考Huiquan WANG 等的方法[17]。
表 1 砂梨花芽采样时间及休眠进程Table 1. Sampling time and dormancy process of P. pyrifolia flower buds黄花梨 Pyrus pyrifolia. Huanghua 蜜雪梨 Pyrus pyrifolia. Mixue 编号
Number采样时间
Sampling date/(年-月-日)休眠进程
Dormancy process编号
Number采样时间
Sampling date/(年-月-日)休眠进程
Dormancy processH1 2013-12-05 未进入休眠
NO dormancyM1 2013-11-29 未进入休眠
No dormancyH2 2013-12-17 内休眠进入
Endodormancy startingM2 2013-12-06 内休眠进入
Endodormancy startingH3 2013-12-25 内休眠
EndodormancyM3 2013-12-12 内休眠
EndodormancyH4 2013-12-31 内休眠
EndodormancyM4 2013-12-19 内休眠
EndodormancyH5 2014-01-08 内休眠解除启动
Endodormancy release startingM5 2013-12-26 内休眠
EndodormancyH6 2014-01-15 内休眠解除
Endodormancy releaseM6 2014-01-04 内休眠
EndodormancyH7 2014-01-21 生态休眠
EcodormancyM7 2014-01-10 内休眠解除启动
Endodormancy release startingH8 2014-01-27 生态休眠
EcodormancyM8 2014-01-16 内休眠解除
Endodormancy releaseH9 2014-02-19 生态休眠
EcodormancyM9 2014-01-21 生态休眠
EcodormancyH10 2014-02-27 花芽萌动
Floral bud germinatingM10 2014-01-26 生态休眠
EcodormancyM11 2014-02-07 花芽萌动
Floral bud germinating1.2 试验方法
1.2.1 内源激素的提取
内源激素的提取参考赵娟娟[18]的方法进行改进:取−80 ℃贮藏的梨花芽0.5 g左右,按质量体积比1∶8加入4 ℃预冷的80%乙醇迅速研磨,收集到10 mL离心管中,4 ℃黑暗条件浸提18 h后,于4 ℃条件下10000 r·min−1离心15 min,取上清液于10 mL离心管中,再按质量体积比1∶5加入预冷的80%乙醇浸提残渣3 h,4 ℃条件下,10000 r·min−1离心15 min,合并上清液,此步骤重复1次。收集得到的上清液倒入旋蒸瓶,蒸干后,加入2 mL色谱甲醇,充分溶解后收集到2 mL离心管中,上清液用C18胶柱去除色素,再用0.22 µm有机系膜过滤,收集于色谱瓶中,待测。
1.2.2 试剂与仪器
激素标样:ABA、GA3、IAA、ZT均购自Sigma公司(美国)。流动相:甲醇、乙腈均为色谱级,购自Merck公司(德国);磷酸缓冲液为pH3.5 PBS。PVPP购自生工生物工程有限公司(上海),甲醇(AR)、乙酸乙酯(AR)、石油醚(AR,沸点:60~90 ℃)均为国产分析纯。超纯水为经0.22 µm水系滤膜过滤双蒸水所得。采用HITACHI(日立)L-2000液相色谱仪,色谱柱为Phenomenex Luna C18 小柱,旋转蒸发仪、0.45 µm有机滤头,0.22 µm水系滤头。
1.3 数据分析
试验数据采用GraphPad Prism 7和Excel软件进行处理。
2. 结果与分析
2.1 黄花梨花芽休眠进程中内源激素的变化
2.1.1 黄花梨花芽休眠进程中内源激素含量的变化
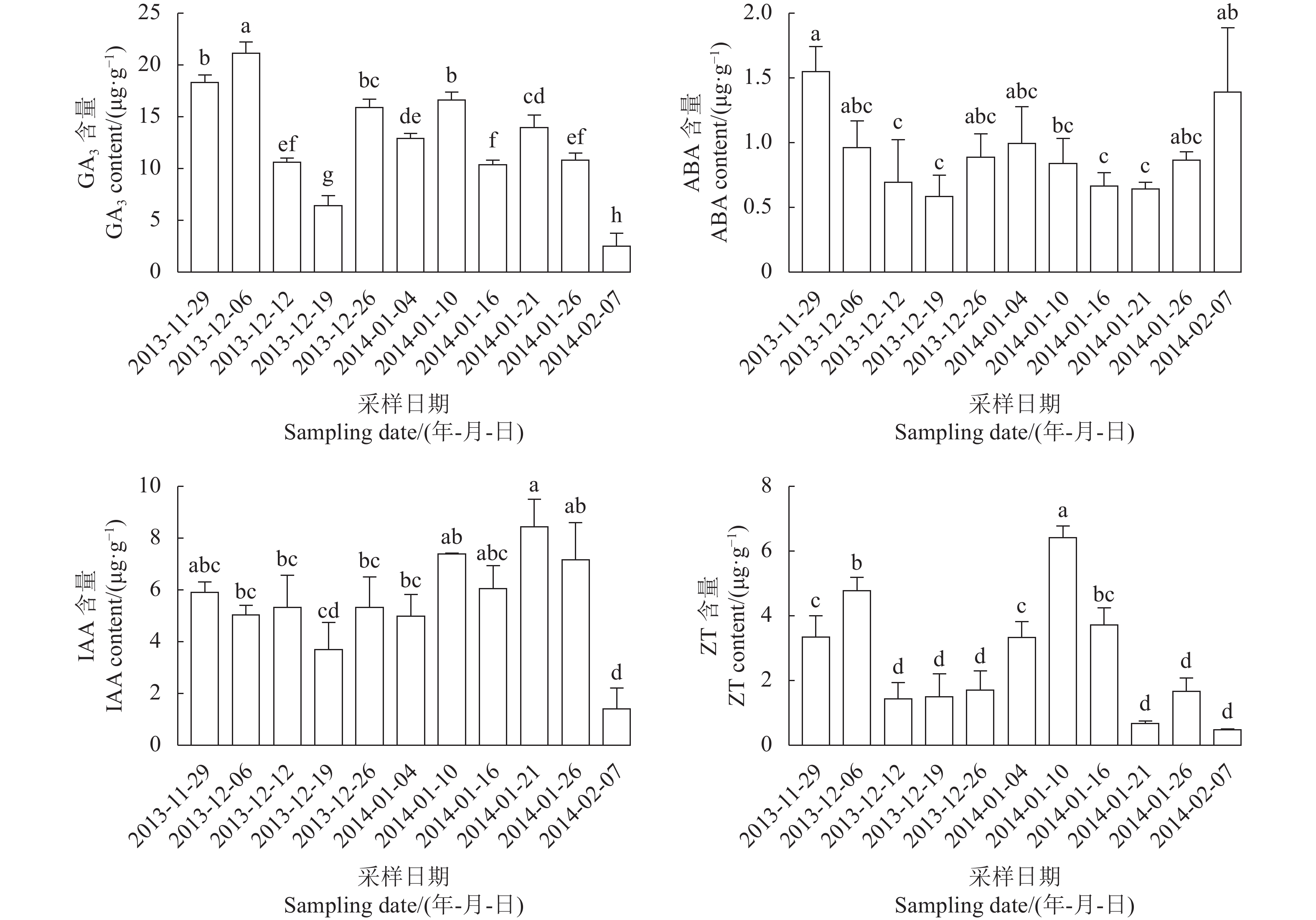
试验结果(图1)可以看出,自2013-12-17(内休眠进入)起GA3含量下降,到2014-01-08(内休眠解除启动)时明显上升,在2014-01-15(内休眠解除)时含量有所下降,在生态休眠期(2014-01-21至2014-02-19)含量有所上升,在2014-02-27(花芽萌动)时含量下降。ABA含量自2013-12-17进入内休眠后呈现了下降的趋势,在2014-01-08(内休眠解除启动)时达到最低值,随后含量有所上升,在2014-01-21(生态休眠期)含量升高,差异显著。IAA含量自2013-12-17(内休眠进入)到2014-01-21(生态休眠期)差异不显著;在2014-01-27(生态休眠期)达到最大值,在2014-02-27(花芽萌动)时含量降到最低。ZT含量在休眠进程中差异显著,自2013-12-17(内休眠进入)到2014-01-08(内休眠解除启动)呈现明显下降趋势,并且在2014-01-08达到最低,在2014-01-15(内休眠解除)后开始上升,在2014-01-21(生态休眠期)含量最高,随后开始下降。
![]()
2.1.2 黄花梨花芽休眠进程中内源激素的平衡关系
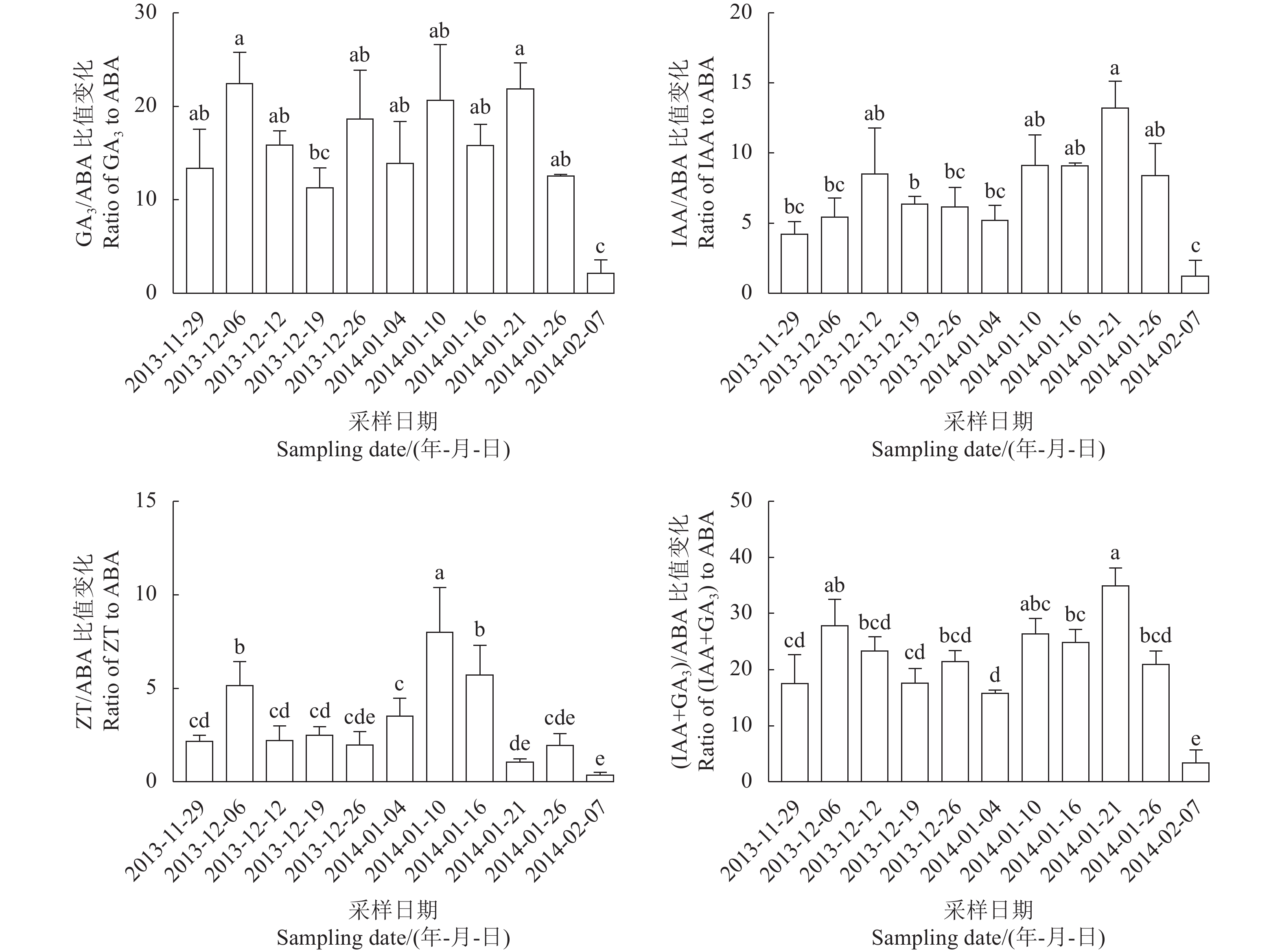
试验结果(图2)可以看出,GA3/ABA比值在2014-01-08(内休眠解除启动)之前均处于较低水平,在内休眠解除启动时比值最大,在2014-01-15(内休眠解除)下降,在2014-01-21下降到最低,随后比值升高,而在2014-02-27(花芽萌动)时达到最低值。IAA/ABA比值除了在2014-01-15(内休眠解除)时出现明显的高峰,在内休眠阶段总体呈上升趋势,只是在2013-12-31出现了下降;在2014-01-21(生态休眠期)出现了明显下降,在2014-02-27(花芽萌动)时比值最低。ZT/ABA比值在2014-01-15(内休眠解除)出现明显高峰,其他时期变化呈现不规律的上下起伏,并且差异显著。(IAA+GA3)/ABA的变化趋势与GA3/ABA比值的变化趋势相同,且各个时期差异显著。在2014-01-8(内休眠解除启动)处于最高水平,在2014-01-15(内休眠解除)时比值略有下降。
![]() 图 2 黄花梨花芽休眠进程中内源激素比值的变化Figure 2. Changes on endogenous hormone ratios in dormancy of Huanghua flower buds
图 2 黄花梨花芽休眠进程中内源激素比值的变化Figure 2. Changes on endogenous hormone ratios in dormancy of Huanghua flower buds2.2 蜜雪梨花芽休眠进程中内源激素的变化
2.2.1 蜜雪梨花芽休眠进程中内源激素含量的变化
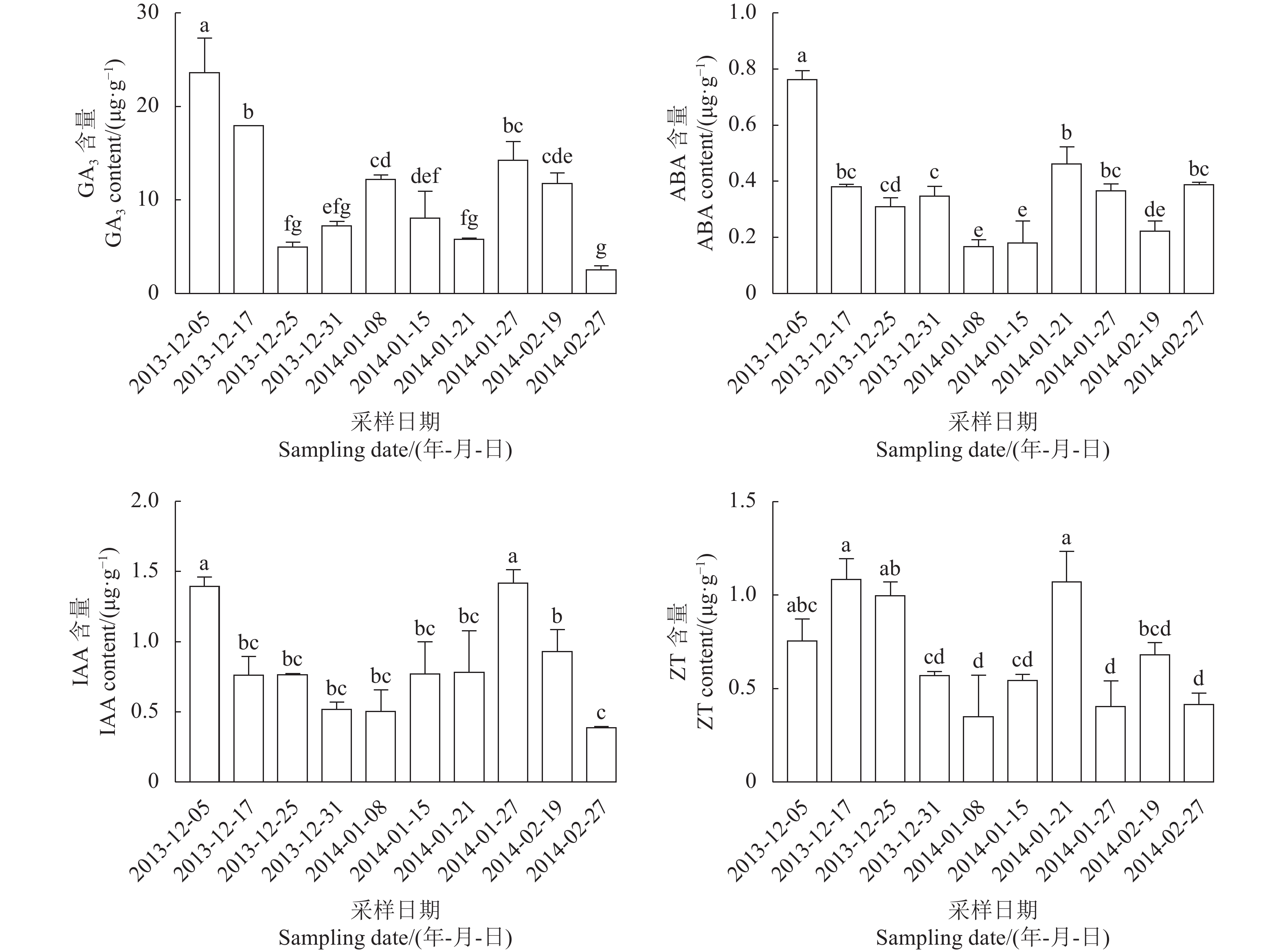
试验结果(图3)可以看出,GA3含量在休眠进程中差异显著。在2013-12-06(内休眠进入)GA3的含量最大,随后在内休眠时期逐渐下降,并在2013-12-19达到较低水平,自2013-12-26开始上升,至2014-01-10(花芽休眠解除启动)时达到峰值,随后又呈现下降趋势,在2014-02-07(花芽萌动)时达到最低水平。ABA含量在2013-11-29最高,在内休眠阶段(2013-12-06至2014-01-04)呈先下降后升高的变化,然后自内休眠解除启动(2014-01-10)起开始下降,至2014-01-21(生态休眠期)再次达到最低值,随后开始升高至2014-02-07(花芽萌动)又达到较高水平,总体呈现明显的“W”型变化趋势。IAA含量自2013-12-06(内休眠进入)至1月4日整个内休眠阶段差异不显著,在2014-01-10(内休眠解除启动)时含量较高,在2014-01-16(内休眠解除)时有所下降,在2014-01-21(生态休眠期)处于最高水平,而在2014-02-07(花芽萌动)时处于最低水平。ZT含量自2013-12-06(内休眠进入)后在内休眠期出现了明显下降,2013-12-12至2013-12-26内休眠时期差异不显著;在2014-01-04开始上升,到2014-01-10(内休眠解除启动)达到最大值,随后在2014-01-16(内休眠解除)时有所下降,并且在生态休眠期均维持在较低水平,并在2014-02-07(花芽萌动)时降到最低水平。
![]() 图 3 蜜雪梨花芽休眠进程中内源激素含量的变化Figure 3. Changes on endogenous hormones in dormancy of Mixue flower buds
图 3 蜜雪梨花芽休眠进程中内源激素含量的变化Figure 3. Changes on endogenous hormones in dormancy of Mixue flower buds2.2.2 蜜雪梨花芽休眠进程中内源激素的平衡关系
从图4可以看出,GA3/ABA、IAA/ABA以及(IAA+GA3)/ABA呈现相似的变化趋势,自2013-12-06(内休眠进入)开始比值呈下降趋势并且均在2014-01-04出现低值,在2014-01-10(内休眠解除启动)升高,而在2014-01-16((内休眠解除)有所下降,在2014-01-21(生态休眠期)达到最大值,随后开始下降,并在2014-02-07(花芽萌动)时达到最低值。ZT/ABA值与其他3个比值变化不同,在2013-12-06(内休眠进入)到2013-12-26内休眠期内ZT/ABA比值差异不显著,无明显变化,在2014-01-10(内休眠解除启动)出现最大值。
![]() 图 4 蜜雪梨花芽休眠进程中内源激素比值的变化Figure 4. Changes on endogenous hormone ratios in dormancy of Mixue flower buds
图 4 蜜雪梨花芽休眠进程中内源激素比值的变化Figure 4. Changes on endogenous hormone ratios in dormancy of Mixue flower buds3. 讨论与结论
植物激素对于植物休眠进程具有调控作用,并且不同激素具有不同的作用[3]。宋猜等[4]、王海波等[5]认为 GA3 是休眠抑制物质,能够代替低温促进内休眠的解除。本研究发现GA3在两个梨品种花芽内休眠进入时含量较高,随着休眠进程含量下降,随后升高,在内休眠解除启动时含量达到较高值,在内休眠解除时又略微下降。这与前人研究结果一致,GA3能够促进梨花芽休眠解除,但本研究显示GA3对于梨花芽的休眠解除作用在于内休眠解除的启动。刘国琴[3]认为GA不是休眠解除的必需因素,而是芽萌发的必需条件,而本研究发现GA3在萌动期含量达到最低,这与本研究结果不一致。
诸多研究表明ABA是休眠的促进物质,对内休眠的解除具有抑制作用。本研究发现ABA的含量在内休眠进入时均处于较高水平,这与高庆玉等[13]研究一致,在芽休眠初期ABA含量最高。黄花梨花芽随着内休眠的进行总体呈现下降的趋势,并在内休眠解除启动时达到最低值,这与牛庆丰[19]、宋猜等[4]认为在内休眠解除前后ABA呈先上升后下降的趋势不同。而在蜜雪梨花芽内休眠进程中呈现先降后升再降的趋势,花芽休眠解除启动时ABA的含量高于休眠解除时的含量。由此可见花芽中ABA含量的降低促进了梨花芽内休眠的解除,但ABA促进休眠解除的作用在不同品种上作用时期有所不同。与其他3种激素相比,ABA的含量较低,这是由于试验采用的是剥鳞的花芽,这与段成国等[14]的结论一致,ABA主要存在于鳞片中。
秦栋等[15]在苹果、马海燕[9]在葡萄上研究发现IAA含量随着自然休眠的解除逐步升高,而本研究发现IAA的含量在两个梨品种花芽内休眠过程中变化差异不明显,说明IAA对于休眠解除的影响较小,这与Sauter等[10]和宋猜等[4]的结论一致,IAA并未直接参与芽的休眠解除。但在本研究中IAA含量在生态休眠期均出现了明显的高峰,推测这与当时环境温度跃变升高有关,温度升高,促进了IAA的合成。但都在萌动期出现了最低值,说明低浓度的IAA促进了芽的萌发,这与段成国等[14]的研究一致,有可能是IAA顶端优势的作用。
段成国等[14]在研究甜樱桃时发现ZT 对内休眠的解除具有促进作用,而本研究发现ZT含量在内休眠进入时含量较高,在内休眠阶段逐渐下降,但在内休眠解除启动期两个梨品种芽内ZT含量正好相反。由此推测ZT对于内休眠解除的调控无直接作用,而是与其他激素共同作用。
内源激素调控植物的休眠并不是单一激素所控制,而是由多种激素之间的平衡所调控[12,20]。本研究通过对各内源激素比值的分析发现,GA3/ABA和(IAA+GA3)/ABA在梨花芽休眠进程中变化趋势基本相同,内休眠解除启动时明显升高,并且在内休眠解除时稍低于内休眠解除启动期,而IAA/ABA比值变化趋势较小,只有在黄花梨花芽中内休眠解除启动时呈现较高的比值。ZT/ABA比值,在两个梨花芽内休眠时期比值变化不规律,但在内休眠解除启动期或内休眠解除期明显升高,这说明ZT/ABA促进了花芽休眠的解除,但是在不同品种中表现出的不同反应。这与丛深等[6]研究葡萄芽休眠时的结果一致,GA3与 ABA 拮抗作用在休眠解除过程中起主导作用,而 ZT、 IAA 和 ABA 拮抗作用则起调节作用。马海燕[9]研究葡萄芽休眠时发现(IAA+GA3)/ABA 比值在休眠解除时逐渐上升,萌芽时达到最大值,这与本研究结果不一致,推测这主要是由花芽萌动时IAA含量极低导致。
本研究对两个砂梨品种花芽休眠进程中內源激素的含量及各内源激素的平衡关系进行了分析,发现GA3和ABA对于休眠起主要作用,ZT不能直接影响花芽休眠进程,而是与ABA共同作用,而IAA对于梨花芽休眠进程影响不大,低浓度的IAA促进了花芽的萌动。 并且不同品种之间內源激素变化不一致,这是因为内源激素本身时一个复杂的系统,同时受到温度变化、不同年份气候、不同水分条件、不同树体营养等方面的影响,不可能呈现完全一致的变化,但是总体有一定的趋势。
-
![]()
![]()
图 2 黄花梨花芽休眠进程中内源激素比值的变化
Figure 2. Changes on endogenous hormone ratios in dormancy of Huanghua flower buds
![]()
图 3 蜜雪梨花芽休眠进程中内源激素含量的变化
Figure 3. Changes on endogenous hormones in dormancy of Mixue flower buds
![]()
图 4 蜜雪梨花芽休眠进程中内源激素比值的变化
Figure 4. Changes on endogenous hormone ratios in dormancy of Mixue flower buds
表 1 砂梨花芽采样时间及休眠进程
Table 1 Sampling time and dormancy process of P. pyrifolia flower buds
黄花梨 Pyrus pyrifolia. Huanghua 蜜雪梨 Pyrus pyrifolia. Mixue 编号
Number采样时间
Sampling date/(年-月-日)休眠进程
Dormancy process编号
Number采样时间
Sampling date/(年-月-日)休眠进程
Dormancy processH1 2013-12-05 未进入休眠
NO dormancyM1 2013-11-29 未进入休眠
No dormancyH2 2013-12-17 内休眠进入
Endodormancy startingM2 2013-12-06 内休眠进入
Endodormancy startingH3 2013-12-25 内休眠
EndodormancyM3 2013-12-12 内休眠
EndodormancyH4 2013-12-31 内休眠
EndodormancyM4 2013-12-19 内休眠
EndodormancyH5 2014-01-08 内休眠解除启动
Endodormancy release startingM5 2013-12-26 内休眠
EndodormancyH6 2014-01-15 内休眠解除
Endodormancy releaseM6 2014-01-04 内休眠
EndodormancyH7 2014-01-21 生态休眠
EcodormancyM7 2014-01-10 内休眠解除启动
Endodormancy release startingH8 2014-01-27 生态休眠
EcodormancyM8 2014-01-16 内休眠解除
Endodormancy releaseH9 2014-02-19 生态休眠
EcodormancyM9 2014-01-21 生态休眠
EcodormancyH10 2014-02-27 花芽萌动
Floral bud germinatingM10 2014-01-26 生态休眠
EcodormancyM11 2014-02-07 花芽萌动
Floral bud germinating 下载: 导出CSV
下载: 导出CSV
-
[1] LIU G Q, LI W S, ZHENG P H, et al. Transcriptomic analysis of ‘Suli’ pear (Pyrus pyrifolia white pear group) buds during the dormancy by RNA-Seq [J]. BMC Genomics, 2012, 13: 700. DOI: 10.1186/1471-2164-13-700
[2] 雷天, 吕招兰. 台农2号蜜雪梨“二次开花”的发生与克服措施 [J]. 果农之友, 2014(2):14,16. DOI: 10.3969/j.issn.1671-7759.2014.02.008 LEI T, LYU Z L. The occurrence and overcoming measures of “second blossoming”of ‘Mixue’ pear [J]. Fruit Growers' Friend, 2014(2): 14,16.(in Chinese) DOI: 10.3969/j.issn.1671-7759.2014.02.008
[3] 刘国琴. 梨休眠分子生理机制研究[D]. 杭州: 浙江大学, 2013. LIU G Q. Studies on molecular mechanism of pear dormancy[D]. Hangzhou: Zhejiang University, 2013. (in Chinese)
[4] 宋猜, 尹明宇, 姜仲茂, 等. ‘优一’杏花芽分化特性及其与休眠期内源激素含量变化的关系 [J]. 西北农林科技大学学报(自然科学版), 2017, 45(6):170−176,184. SONG C, YIN M Y, JIANG Z M, et al. Relationship between flower bud differentiation and change of endogenous hormone of ‘Youyi’ during dormancy period [J]. Journal of Northwest A & F University (Natural Science Edition), 2017, 45(6): 170−176,184.(in Chinese)
[5] 王海波, 高东升, 王孝娣, 等. 赤霉素和脱落酸与桃芽自然休眠诱导 [J]. 果树学报, 2006, 23(4):599−601. DOI: 10.3969/j.issn.1009-9980.2006.04.024 WANG H B, GAO D S, WANG X D, et al. Role of gibberellin and abscisic acid in peach bud endodormancy induction [J]. Journal of Fruit Science, 2006, 23(4): 599−601.(in Chinese) DOI: 10.3969/j.issn.1009-9980.2006.04.024
[6] 丛深, 王海波, 王孝娣, 等. 带叶休眠对休眠解除期间葡萄芽呼吸代谢的影响 [J]. 园艺学报, 2013, 40(10):1983−1989. DOI: 10.3969/j.issn.0513-353X.2013.10.014 CONG S, WANG H B, WANG X D, et al. Effect of dormancy with leaves on respiration of grape buds during dormancy release [J]. Acta Horticulturae Sinica, 2013, 40(10): 1983−1989.(in Chinese) DOI: 10.3969/j.issn.0513-353X.2013.10.014
[7] 孙利鑫, 张亚红. 落叶果树休眠期解除界定的研究进展 [J]. 中国果树, 2016(2):55−59. DOI: 10.16626/j.cnki.issn1000-8047.2016.02.014 SUN L X, ZHANG Y H. Research progress on the definition of dormancy release of deciduous fruit trees [J]. China Fruits, 2016(2): 55−59.(in Chinese) DOI: 10.16626/j.cnki.issn1000-8047.2016.02.014
[8] 郑国生, 盖树鹏, 盖伟玲. 低温解除牡丹芽休眠进程中内源激素的变化 [J]. 林业科学, 2009, 45(2):48−52. DOI: 10.3321/j.issn:1001-7488.2009.02.009 ZHENG G S, GAI S P, GAI W L. Changes of endogenous hormones during dormancy release by chilling in tree peony [J]. Scientia Silvae Sinicae, 2009, 45(2): 48−52.(in Chinese) DOI: 10.3321/j.issn:1001-7488.2009.02.009
[9] 马海燕. 葡萄生长过程中内源激素含量变化的研究[D]. 杨凌: 西北农林科技大学, 2007. MA H Y. Changes of endogenous hormones in grapevine during its development[D]. Yangling: Northwest A & F University, 2007. (in Chinese)
[10] SAUTER A, ABRAMS S R, HARTUNG W. Structural requirements of abscisic acid (ABA) and its impact on water flow during radial transport of ABA analogues through maize roots [J]. Journal of Plant Growth Regulation, 2002, 21(1): 50−59. DOI: 10.1007/s003440010040
[11] 毕磊. 七月酥梨休眠期内含物变化及生理代谢研究[D]. 保定: 河北农业大学, 2009. BI L. Study on changes of inclusions and metabolism during dormant period of qiyuesu pear[D]. Baoding: Hebei Agricultural University, 2009. (in Chinese)
[12] 袁燕波, 王历慧, 于晓南. 芍药休眠芽发育进程内源激素变化研究 [J]. 浙江农业学报, 2014, 26(1):54−60. DOI: 10.3969/j.issn.1004-1524.2014.01.10 YUAN Y B, WANG L H, YU X N. Comparative analysis of endogenous hormones during dormant bud development of Chinese herbaceous peony [J]. Acta Agriculturae Zhejiangensis, 2014, 26(1): 54−60.(in Chinese) DOI: 10.3969/j.issn.1004-1524.2014.01.10
[13] 高庆玉, 闫超, 张丙秀, 等. 树莓果实与休眠芽生长过程中内源激素含量变化研究 [J]. 东北农业大学学报, 2015, 46(1):19−25. DOI: 10.3969/j.issn.1005-9369.2015.01.004 GAO Q Y, YAN C, ZHANG B X, et al. Study on the variation of endogenous hormone content during the growth of raspberry fruit and dormant bud [J]. Journal of Northeast Agricultural University, 2015, 46(1): 19−25.(in Chinese) DOI: 10.3969/j.issn.1005-9369.2015.01.004
[14] 段成国, 李宪利, 高东升, 刘焕芳, 李萌. 剥鳞和激素处理对大樱桃花芽休眠解除及内源激素变化的影响 [J]. 西北植物学报, 2004, 24(4):615−620. DOI: 10.3321/j.issn:1000-4025.2004.04.009 DUAN C G, LI X L, GAO D S, et al. Effects of removing scales and exogenous hormone treatments on changes of endogenous hormone in sweet cherry flower buds and dormancy release during dormancy [J]. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(4): 615−620.(in Chinese) DOI: 10.3321/j.issn:1000-4025.2004.04.009
[15] 秦栋. 酸王(Malus domestica cv.‘Avrolles’)自然休眠解除后生理生化变化研究[D]. 泰安: 山东农业大学, 2009. QING D. Study on the physiological and biochemical changes in Avrolles(Malus domestica)after natural dormancy releasing[D]. Taian: Shandong Agricultural University, 2009. (in Chinese)
[16] 刘冬峰, 陈露露, 李国鹏, 等. 氰胺对‘翠冠’梨花芽休眠解除及内源激素和碳水化合物含量的影响 [J]. 浙江大学学报(农业与生命科学版), 2013, 39(1):69−74. LIU D F, CHEN L L, LI G P, et al. Effect of hydrogen cyanamide(HC) on release of floral bud dormancy, endogenous hormone and carbohydrate in Pyrus pyrifolia cv. Cuiguan [J]. Journal of Zhejiang University (Agriculture and Life Sciences), 2013, 39(1): 69−74.(in Chinese)
[17] WANG H Q, LIN Z Y, CHEN J N, et al. Classification of dormancy stage in Asian pear buds under the influence of, natural environment [J]. Fresenius Environmental Bulletin, 2020, 29(2): 1241−1249.
[18] 赵娟娟. 柚果实生长过程中内源激素含量变化的研究[D]. 福州: 福建农林大学, 2013 ZHAO J J. Study on the changes of endogenous hormone contents in pomelo fruit growth process[D]. Fuzhou: Fujian Agriculture and Forestry University, 2013. (in Chinese)
[19] 牛庆丰. 梨花芽休眠转换的分子网络及调控机制[D]. 杭州: 浙江大学, 2016. NIU Q F. The molecular mechanism and molecular network of pear flower bud dormancy transition[D]. Hangzhou: Zhejiang University, 2016. (in Chinese)
[20] SEELEY S D, DAMAVANDY H, ANDERSON J L, et al. Autumn-applied growth regulators influence leaf retention, bud hardiness, bud and flower size, and endodormancy in peach and cherry [J]. Journal of the American Society for Horticultural Science, 1992, 117(2): 203−208. DOI: 10.21273/JASHS.117.2.203
-
期刊类型引用(2)
1. 盛盼进,魏希,张北红,另青艳,胡锦,肖祖飞. 6-BA对芳樟矮林萌芽及生长的影响. 安徽农学通报. 2023(17): 86-89 .  百度学术
百度学术
2. 朱璟. 农业资源税对农业产业转型升级的促进作用. 黑河学院学报. 2023(09): 50-52 . 百度学术
其他类型引用(3)

计量
- 文章访问数: 711
- HTML全文浏览量: 295
- PDF下载量: 59
- 被引次数: 5
