

Effects of Drying Technology on Nutrition of Tremella fuciformis
-
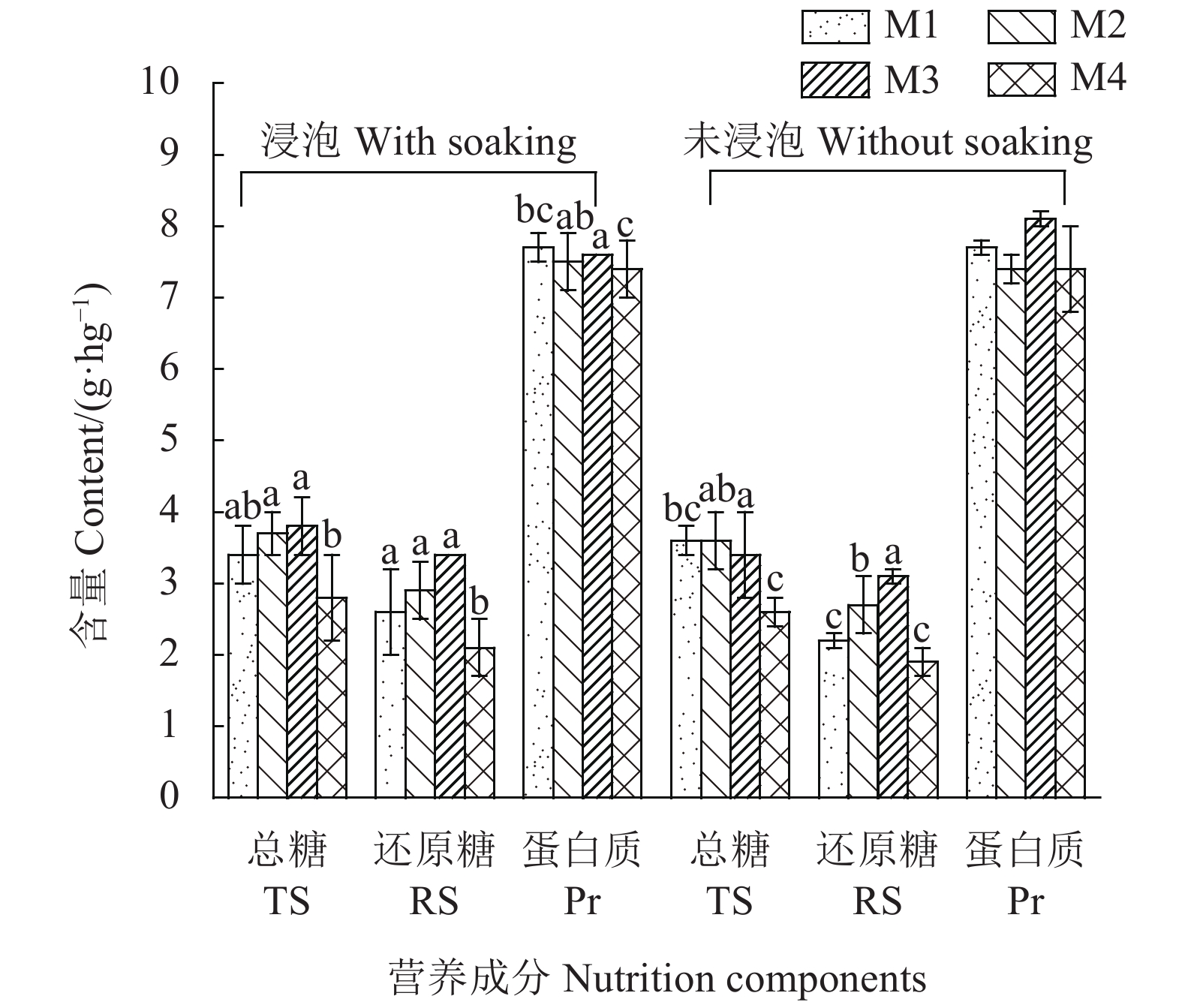
摘要:目的 研究烘干方式对银耳营养的影响,为完善银耳烘干工艺、提高银耳品质提供理论依据。方法 以蛋白质、糖类含量和氨基酸营养品质为评价指标,应用统计学和FAO/WHO建议的氨基酸评估模式,比较银耳实际生产中常见的以废菌棒为燃料工厂化烘干(M1)、以废菌棒木屑为燃料工厂化烘干(M2)、以废菌棒燃料小灶烘干(M3)、电热工厂化烘干(M4)等4种烘干工艺对银耳营养的影响。结果 烘干前的浸泡工序对银耳蛋白质、糖类、氨基酸营养和二氧化硫含量无显著性影响(P>0.05)。但M2、M3烘干条件下的银耳总糖、还原糖含量显著高于M4(P<0.05)。4种烘干方式制备的银耳干品单一氨基酸含量及总量均无显著性差异(P>0.05),组成特征相似。不同烘干组银耳氨基酸营养平衡性略有差异,氨基酸比值系数分从高到低依次为M1(80.52)>M4(79.55)>M3(79.15)>M2(77.80),说明银耳是一种氨基酸营养比较均衡的食物。赖氨酸为银耳干制品的第一限制性氨基酸。结论 烘干前的清洗、浸泡不会导致银耳营养变化,现行烘干工艺制备的银耳干品并无二氧化硫残留,以废菌棒木屑为燃料的工厂化烘干方式及以废菌棒燃料的小灶烘干方式下银耳营养损失较少。Abstract:Objective Effect of drying technology on the nutritional quality of Tremella fuciformis Berk was studied.Method The national standards on the nutritional quality of edible fungi, such as contents of total sugar, reducing sugar, protein, and amino acids, were the indices used to evaluate 4 drying technologies in producing the dehydrated product. The FAO/WHO indexing system was applied to gage the protein nutrition. Dehydration of T. fuciformis employed either spent culture substrate for fuel in a factory oven (M1), spent culture substrate and wood chips for fuel in a factory oven (M2), spent culture substrate for fuel in a workshop oven (M3), or an industrial electric oven (M4).Results Water-soaking of the fungi prior to dehydration showed no significant effect on the contents of protein, sugar, amino acids, and sulfur dioxide in the dried product (P>0.05). However, the various drying methods resulted in quality differentiations among them. The contents of total sugar and reducing sugar in the dried T. fuciformis prepared under M2 and M3 were significantly higher than those under M4 (P<0.05). The amino acid contents of the products made by different technologies did not differ significantly (P>0.05). The nutritional grading based on amino acids ranked the dried mushrooms from those made by M1 at a score of 80.52, to M4 of 79.55, M3 of 79.15, and M2 of 77.80. All dried fungi products had a well-balance nutritional composition with lysine being the top restricted amino acid.Conclusion The total sugar and amino acids in the dehydrated T. fuciformis were not affected by the pre-soaking but significantly by the drying process. Either drying by using spent culture substrate and wood chips for fuel in a factory oven (M2) or by using spent culture substrate for fuel in a workshop oven (M3) retained the nutrients more effectively than other methods. Incidentally, there was no sulfur dioxide residue detected in the dried T. fuciformis produced by the process currently practiced by the industry.
-
Keywords:
- Tremella fuciformis Berk /
- drying /
- nutrition property /
- amino acid /
- sulfur dioxide
-
0. 引言
【研究意义】百香果(Passiflora edulis Sims)属西番莲科(Passifloraceae)西番莲属(Passiflora)植物,是广西的特色水果之一。茎基腐病是百香果主要真菌病害之一,其病原为百香果茎基腐病菌(Fusarium solani),该病发生在植株茎基部,与地面有20~30 cm的距离,幼苗、成株均会被病菌侵染,造成植株褪绿萎蔫或枯萎死亡[1],严重影响百香果的产量和品质。目前百香果茎基病的生产防治技术还是以化学防治为主[2]。化学防治虽简便快速,但化学药剂长期使用易造成环境污染、生态失衡等问题,甚至威胁人类生命安全,因此,植物病害生物防治被认为是一种更安全和可持续发展的战略[3]。生物防治是推动农业可持续发展的重要组成,其借助自然界中不影响植物生长的微生物发挥其重寄生、竞争等多种生防作用,以直接或间接作用的方式影响病原物的生存或活动;同时可诱导植物的抗病性或促进植株生长,从而达到保护植株的目的[4]。与化学防治药剂相比,拮抗菌产生的抑菌物质具有毒性低、对环境友好,降低病原微生物耐药性的优点,可安全地应用于植物病害防治[5]。但菌株在实际应用过程中,其在土壤中的稳定性和在植物中的定植能力问题有待进一步研究和解决[6],才能提高菌株的存活和增殖能力,确保菌株在可持续农业中的商业化应用推广。【前人研究进展】Wang等[7]从土壤中分离得到贝莱斯芽胞杆菌(Bacillus velezensis)HNAH
17806 ,并从其发酵液中纯化出一种分子量约为 1.5 kDa 的抗真菌肽(BVAP),对菌丝抑制的 IC50 为 5.58 μg·mL−1,其作用机制为增加菌丝的通透性、导致菌丝尖端肿胀。Chen 等对分别从香蕉和茶树根际土壤中分离得到的枯草芽胞杆菌 YBC 和 151B1 进行研究,结果表明菌株在 SYB 培养基的无菌发酵液影响了 F. solani 细胞膜的完整性、抑制分生孢子的萌发和菌丝生长[8]。这些研究表明,拮抗菌在防治百香果茎基腐病中展现了广泛的应用前景,为研究拮抗菌与病原菌互作机制提供了理论参考。拮抗菌通过产生胞外酶水解病原菌细胞壁以破坏菌丝,抑制菌丝生长,常见的细胞壁降解酶类主要有纤维素酶、蛋白酶、几丁质酶、β-1,3-葡聚糖酶等[9]。除了直接抑制病原菌,拮抗菌还表现出显著的植物促生作用。例如,贝莱斯芽胞杆菌W-1对花椒根腐病的抑菌率高达89%,其通过生物固氮、磷酸盐溶解以及分泌植物激素如吲哚-3-乙酸(IAA)、细胞分裂素(CTK)和赤霉素(GA),有效促进植物生长[10,11],为芽胞杆菌在多种作物病害防治中的应用提供了理论支持。此外,杨东亚等[12]发现菌株XY-1、XY-13、XY-53不仅对腐皮镰刀菌抑制效果显著,还能通过产蛋白酶、固氮等途径,显著提高黄瓜幼苗的生物量,进一步证实了拮抗菌在病害防治和植物促生中的双重作用。油茶根部内生的芽胞杆菌AF9和AH6则被发现具有诱导植物产生系统抗性(ISR)的能力[13]。李丽丽等[13]研究表明,这些菌株不仅能有效降低油茶根腐病的发生,还能通过增强植物的抗病性和促进生长,展现出广泛的生防潜力。【本研究切入点】尽管前人研究已经表明芽胞杆菌在植物病害生物防治中具有显著的潜力,但关于百香果茎基腐病的生物防治研究较少,特别是在高效生防菌株的筛选和其作用机制方面的研究还不够深入。【拟解决的关键问题】从健康的百香果根际土壤中筛选鉴定拮抗细菌,并进行初步的生防、促生作用研究,进一步明确拮抗菌与病原菌的作用机制,以期为百香果茎基腐病生物防治提供新的菌种资源和理论依据,最终推动生防制剂的开发和应用。1. 材料与方法
1.1 试验材料
1.1.1 培养基
马铃薯葡萄糖琼脂培养基(potato dextrose agar,PDA)[14]:葡萄糖20 g、琼脂20 g 、土豆粉6 g,去离子水定容至1 000 mL。
LB培养基(luria-bertani medium)[15]:胰蛋白胨10 g、酵母提取物5 g、NaCl 10 g。(LB固体培养基在LB培养基的基础上加入18 g琼脂)。
1.1.2 根际土壤样品采集
广西南宁市上林县白圩镇(108°48′ 28.54″E, 23°26′ 17.25″N)健康黄金、维蜜、钦蜜百香果根际土各2份,广西南宁市良庆区那马镇(108°26′ 43.13″E,22°38′ 40.82″N)健康台农一号百香果根系土1份。
1.2 试验方法
1.2.1 根际土壤细菌分离
采用稀释涂布法[16]分离土壤中的细菌,称取1 g根际土壤样品,加入9 mL无菌水,并稀释至10−5倍液,分别吸取10−3、10−4、10−5倍液各100 µL涂板,每个浓度重复3次,置于28 ℃培养箱中培养。根据菌落大小、颜色、质地、光泽及表面形态等特征挑选形态差异显著的单菌落用平板划线法纯化,转接至新的LB固体培养基平板上培养,4 ℃保存备用。
1.2.2 拮抗细菌的筛选
采用平板对峙法对分离纯化的菌株进行初筛和复筛,以F. solani为靶标菌,当菌落直径培养至6~7 cm后,从菌丝边缘打取菌饼接种至PDA培养基中央,并在距病原菌2.2 cm的四周接种4种不同的待测菌株,对照为只接种病原菌,设3次重复,置于28 ℃培养箱中培养,待对照菌丝长满平板,测量各处理的菌落直径大小,计算抑制率 [17]。抑制率/%=[(对照菌落直径−菌饼直径)−(处理菌落直径−菌饼直径)]/(对照菌落直径−菌饼直径)×100。将初筛具有明显抑制作用的菌株进行复筛,在距病原菌2.2 cm的两侧接种同种菌株,其余步骤参照初筛。
1.2.3 拮抗菌鉴定
将菌株划线培养于LB固体培养基后置于28 ℃培养箱中培养5 d 后观察不同菌株单菌落的形态特征。采用菌落PCR的方法,以通用引物27F/1492R[18]进行PCR扩增,PCR反应体系(50.0 μL):2×Taq MasterMix 25.0 μL,上、下游引物各2.0 μL,DNA模板为少量菌体,ddH2O 19.0 μL;反应程序:预变性95 ℃、5 min;变性95 ℃、30 s,退火56 ℃、30 s,延伸72 ℃、1 min,33个循环;终延伸72 ℃、10 min。扩增产物纯化回收,并克隆至载体pUC-19T上(具体步骤参照Takara TA克隆试剂盒说明书),送至上海生工生物有限公司进行测序,并将测序结果在National Center for Biotechnology Information(https://www.ncbi.nlm.nih.gov/BLAST/)数据库中进行比对,利用 MAGA11.0软件中的邻接法构建菌株与其相关近缘属种的系统发育树。
1.2.4 拮抗菌抗生素相关基因检测
参照宋文欣[19]的引物进行抗生素基因引物合成。以拮抗菌株的总DNA为模板,以表1所示引物进行PCR扩增,对mycB(抗霉枯草菌素)、ifuA(伊枯草菌素)、yndj、fenB(丰原素)和bacA(溶杆菌素)5种抗生素产生相关基因进行检测,扩增产物经1.0%琼脂糖凝胶电泳检测。
表 1 检测抗生素产生相关基因的引物序列Table 1. Primers for detecting genes related to antibiotic production基因名称
Genes引物名称
Primer引物序列(5′-3′)
Primer sequences退火温度
Annealing temperature/ ℃mycB MycB-F
MycB-RATGTCGGTGTTTAAAAATCAAGTAACG
TTAGGACGCCAGCAGTTCTTCTATTGA55 fenB FenB-F
FenB-RCTATAGTTTGTTGACGGCTC
CAGCACTGGTTCTTGTCGCA55 ifuA IfuA-F
IfuA-RATGTATACCAGTCAATTCC
GATCCGAAGCTGACAATAG55 bacA BacA-F
BacA-RCAGCTCATGGGAATGCTTTT
CTCGGTCCTGAAGGGACAAG58 yndj 147-F
147-RCAGAGCGACAGCAATCACAT
TGAATTTCGGTCCGCTTATC55 1.2.5 拮抗菌胞外酶检测
参考宋文欣[19]的方法,将拮抗菌株点接至10 mL LB培养基置于28 ℃、200 r·min−1的摇床培养12 h后吸取500 μL至新的20 mL LB培养基中扩大培养至吸光度D600=0.6,吸取菌液1 μL均匀的点接在距蛋白酶、纤维素酶、几丁质酶、β,1-3-葡聚糖酶、木聚糖酶检测平板中心2.2 cm处,CK为点接等量的LB培养基,重复3次,置于28 ℃培养箱中培养3 d,观察菌落周围是否产生透明圈,若产生透明圈,测量菌株直径和透明圈直径的大小。而纤维素酶活性检测在培养相同时间后加入10 mL的1 mg·mL−1刚果红静置30 min后,倒掉该溶液,加入等量的1 mol·L−1 NaCl静置30 min后倒掉,观察菌落周围是否产生透明圈,若产生透明圈,测量菌株及透明圈直径的大小。
1.2.6 拮抗细菌菌株对番茄种子的促生作用研究
参考杨晓云等[20]的方法并稍改动,先用无菌水浸泡番茄的种子(品种为抗枯番茄)12 h,接着用1%次氯酸钠溶液浸泡消毒10 min,再用无菌水清洗3次后置于无菌培养皿中待用;将拮抗菌培养至吸光度D600=0.8,吸取5 mL菌液浸泡12 h,对照为等量的LB培养基,经菌液浸泡过的种子用无菌水清洗表面菌体,吸干表面水分,将种子置于有湿润滤纸的瓶子中,每处理20颗,设3次重复。48 h记录种子的发芽率,7 d后测量种子的胚根长、胚芽长等生物量,计算种子的平均发芽率,胚根和胚芽的均值和标准差。
1.2.7 拮抗菌相容性研究
于同一LB固体培养基平板上接种不同菌株研究其相容性,将拮抗细菌划线接种至LB固体培养基上并两两相交,置于28 ℃培养箱中培养5 d,试验重复3次,观察菌株间是否存在抑菌带或抑菌圈,上述两种现象均无表示菌株相容,反之则不相容[21]。
1.3 数据统计与分析
使用软件IBM SPSS Statistics 25进行统计学分析,采用Duncan新复极差法进行多重比较。
2. 结果与分析
2.1 拮抗菌分离与筛选
采用平板对峙法对分离纯化菌株进行筛选,共分离得到52株菌株,其中8株具有拮抗作用,对病原菌的抑制率介于42.18%~57.81%(图1),其中NN-Q01-10、SL-Q02-05、SL-Q02-06对病原菌抑制效果最好,抑制率分别为57.81%、57.34%、57.34%。
![]() 图 1 拮抗菌株对F. solani平板抑制效果Figure 1. Antagonistic effect on F. solani plate inhibition shown by isolated strains
图 1 拮抗菌株对F. solani平板抑制效果Figure 1. Antagonistic effect on F. solani plate inhibition shown by isolated strains2.2 拮抗菌鉴定
2.2.1 形态学观察
拮抗细菌形态学观察结果(图2)显示SL-Q02-04、SL-Q02-06、SL-Q03-01、NN-Q04-01菌落近圆形,表面干燥,易挑起,不凸起,表面有少量杆状褶皱,菌落米白色,边缘不光滑,不透明。SL-Q02-05、NN-Q03-01菌落圆形,表面干燥,易挑起,不凸起,表面有杆状褶皱,质地较粗糙,菌落米白色,边缘不光滑,不透明。NN-Q03-09菌落圆形,表面干燥,凸起,易挑起,有褶皱,菌落白色,边缘不光滑,不透明。NN-Q03-10菌落圆形,表面干燥,凸起,不易挑起,有褶皱,菌落白色,边缘不光滑,不透明。
![]() 图 2 拮抗菌形态学观察A:拮抗菌在LB固体培养基平板上的整体形态图;B:拮抗菌在LB固体培养基平板上的菌落形态图 。Figure 2. Morphology of antagonistic bacteriaA: The overall morphology of antagonistic bacteria on LB solid medium plates B: Colony morphology of antagonistic bacteria on LB solid medium agar plates.
图 2 拮抗菌形态学观察A:拮抗菌在LB固体培养基平板上的整体形态图;B:拮抗菌在LB固体培养基平板上的菌落形态图 。Figure 2. Morphology of antagonistic bacteriaA: The overall morphology of antagonistic bacteria on LB solid medium plates B: Colony morphology of antagonistic bacteria on LB solid medium agar plates.2.2.2 分子鉴定
分别以8个拮抗菌株的DNA为模板扩增16S rDNA,得到
1514 bp大小的片段,测序结果提交至NCBI获得登录号(PP917510~PP917517)。在NCBI中进行Blast比对,并基于16S rDNA基因构建进化树,以蜡样芽胞杆菌(B. cereus)为外群(图3)。其中,SL-Q02-04与Bacillus subtilis subsp. subtilis str. 168聚为一支,SL-Q02-06、SL-Q03-01与Bacillus subtilis strain QB61聚为一支,NN-Q04-01与Bacillus inaquosorum strain BIM B-2002聚为一支,SL-Q02-05、NN-Q03-01、NN-Q03-09、NN-Q03-10与Bacillus velezensis strain XHA16聚为一支,表明分离菌株主要属枯草芽胞杆菌(B. subtilis)、贝莱斯芽胞杆菌(B. velezensi)、枯草芽胞杆菌沙漠亚种(B. subtilis subsp. inaquosorum)。![]() 图 3 基于16S rDNA基因序列构建的拮抗菌及相关近缘种的系统发育树Figure 3. Phylogenetic tree of antagonistic bacteria and related species based on 16S rDNA gene sequences
图 3 基于16S rDNA基因序列构建的拮抗菌及相关近缘种的系统发育树Figure 3. Phylogenetic tree of antagonistic bacteria and related species based on 16S rDNA gene sequences2.3 拮抗菌抗生素相关基因检测
由抗生素合成相关基因PCR产物电泳结果(图4)可以看出8个菌株均具有bacA、FenB、yanJ抗生素基因,推测其基因组具有可产生相应的抗生素类物质的基因,此外,SL-Q02-05、NN-Q03-01、NN-Q03-09、NN-Q03-10扩增得到MycB(抗霉枯草菌素)、ifuA(伊枯草菌素)相应的单一条带。根据抗生素相关基因检测结果推测拮抗菌株可能在发酵培养条件下产生相关抗生素,可进一步进行抗生素合成基因簇预测,及抗生素提取、纯化、鉴定等试验验证。
![]() 图 4 抗生素合成基因PCR产物Figure 4. Electrophoresis of PCR products of antibiotics synthesis genesM: Marker DL2000; 1: SL-Q02-04; 2: SL-Q02-05; 3: SL-Q02-06; 4: SL-Q03-01; 5: NN-Q03-01; 6: NN-Q03-09; 7: NN-Q03-10; 8: NN-Q04-01.
图 4 抗生素合成基因PCR产物Figure 4. Electrophoresis of PCR products of antibiotics synthesis genesM: Marker DL2000; 1: SL-Q02-04; 2: SL-Q02-05; 3: SL-Q02-06; 4: SL-Q03-01; 5: NN-Q03-01; 6: NN-Q03-09; 7: NN-Q03-10; 8: NN-Q04-01.2.4 拮抗细菌胞外酶活性
通过蛋白酶、纤维素酶、β-1,3-葡聚糖酶及木聚糖酶的检测平板测定8个拮抗菌株的胞外酶活性。结果显示,所有菌株均可分泌4种胞外酶,NN-Q03-09产蛋白酶能力最强,同时拮抗菌株在β-1,3-葡聚糖酶检测平板上产生的透明圈边界不清晰,证明产生β-1,3-葡聚糖酶的能力较弱,而在木聚糖平板上可产生相对明显的透明圈(图5)。
![]() 图 5 拮抗菌胞外酶测定从上至下依次为蛋白酶、纤维素酶、β-1,3-葡聚糖酶、木聚糖酶。Figure 5. Assay on antimicrobial extracellular enzymesFrom top to bottom: protease, cellulase, β-1,3-glucanase, and xylanase.
图 5 拮抗菌胞外酶测定从上至下依次为蛋白酶、纤维素酶、β-1,3-葡聚糖酶、木聚糖酶。Figure 5. Assay on antimicrobial extracellular enzymesFrom top to bottom: protease, cellulase, β-1,3-glucanase, and xylanase.2.5 拮抗细菌菌株对番茄种子促生作用研究
番茄种子在不同拮抗菌液处理后,结果显示番茄种子的长势较好,有6个菌株(SL-Q02-04、SL-Q02-05、SL-Q02-06、SL-Q03-01、NN-Q03-01、NN-Q03-10)对胚根生长具有明显的促进作用,其中SL-Q02-04最为明显,该菌株菌液处理后番茄幼苗的胚根最长为7.32 cm;所有菌株对番茄发芽率和胚芽生长均没有促进作用(图6、表2)。
![]() 图 6 不同拮抗菌处理下番茄幼苗(7 d)Figure 6. Tomato seedlings under antagonistic treatments for 7 d表 2 不同拮抗菌处理下番茄种子的生物量统计Table 2. Statistics on tomato seed biomass under antagonistic treatments
图 6 不同拮抗菌处理下番茄幼苗(7 d)Figure 6. Tomato seedlings under antagonistic treatments for 7 d表 2 不同拮抗菌处理下番茄种子的生物量统计Table 2. Statistics on tomato seed biomass under antagonistic treatments处理
Treatment发芽率
Germination percentage/%胚根长
Embryo root
length/cm胚芽长
Embryo length/cmCK 0.87±0.03a 4.76±0.24d 5.31±0.11a SL-Q02-04 0.75±0.00abc 7.32±0.06a 5.39±0.61a SL-Q02-05 0.83±0.07ab 6.03±0.04bc 5.77±0.17a SL-Q02-06 0.72±0.02bc 6.65±0.12ab 5.16±0.33a SL-Q03-01 0.87±0.08abc 6.17±0.24b 5.83±0.10a NN-Q03-01 0.77±0.07c 6.67±0.14ab 4.50±0.13a NN-Q03-09 0.85±0.03ab 5.07±0.51d 4.70±1.02a NN-Q03-10 0.87±0.05a 6.63±0.16ab 5.83±0.25a NN-Q04-01 0.77±0.02abc 5.29±0.21cd 5.33±0.11a 表中数据为平均数±标准误。同列不同小写字母表示经Duncan氏新复极差法检验差异显著(P<0.05)。
Data are average ± standard error; those with different lowercase letters on same column indicate significant difference by Duncan's new complex range test (P<0.05).2.6 拮抗菌相容性
对8个拮抗菌株进行相容性测定,结果显示菌株接种培养5 d后(图7),NN-Q04-01与剩余菌株均不相容,SL-Q02-05、SL-Q02-06、SL-Q03-01、NN-Q03-09间不相容,SL-Q02-06与NN-Q03-01不相容,SL-Q03-01与NN-Q03-01不相容,部分相容性较好的拮抗菌株可用于后续的复合使用。
![]() 图 7 拮抗菌相容性Figure 7. Compatibility between antagonistic bacteria1: SL-Q02-04; 2: SL-Q02-05; 3: SL-Q02-06; 4: SL-Q03-01; 5: NN-Q03-01; 6: NN-Q03-09; 7: NN-Q03-10; 8: NN-Q04-01.
图 7 拮抗菌相容性Figure 7. Compatibility between antagonistic bacteria1: SL-Q02-04; 2: SL-Q02-05; 3: SL-Q02-06; 4: SL-Q03-01; 5: NN-Q03-01; 6: NN-Q03-09; 7: NN-Q03-10; 8: NN-Q04-01.3. 讨论与结论
生物防治是农业生产上安全、有效的防控方法,近年来研究应用较为广泛。然而,关于百香果茎基腐病的生物防治研究相对较少,主要以化学防治为主,尽管已有部分拮抗真菌和细菌被筛选出来,但其应用范围仍然有限。菌株筛选和鉴定是拮抗菌开发和应用的基础[22],本研究从健康的百香果根际土壤分离筛选出8株具有抑制作用的拮抗菌株,主要包括枯草芽胞杆菌、贝莱斯芽胞杆菌、枯草芽胞杆菌沙漠亚种。这表明健康植株根际土壤是有效生防材料的来源之一。
Wang等[7]从贝莱斯芽胞杆菌B. velezensis HNAH
17806 发酵液中分离得到抗真菌肽BVAP,并鉴定为丰原素,其通过增加真菌膜通透性,诱导菌丝膨胀,导致核酸和几丁质在膨胀部位异常积累。本研究中贝莱斯芽胞杆菌B. velezensis菌株NN-Q03-01、NN-Q03-09、NN-Q03-10可检测到FenB抗生素基因,这与Wang等[7]的研究结果一致,为后续进一步验证该抗生素基因的功能提供了基础。Chen等[8]研究发现枯草芽胞杆菌菌株151B1和YBC的发酵菌液可影响腐皮镰刀菌F. solani的膜完整性并诱导其凋亡。同时151B1菌株还能产生iturin A化合物,推测该脂肽可诱导病原菌的凋亡。此外,Ntushelo等[23]指出,iturins与真菌细胞膜接触后可形成离子传导孔,破坏质膜,导致细胞内含物渗漏。本研究发现,枯草芽胞杆菌SL-Q02-05能够扩增出ifuA抗生素基因,与前人研究结果一致,后续研究将进一步验证该菌株的抗菌活性。植物病原菌细胞壁主要由纤维素、葡聚糖、蛋白质组成,胞外酶的产生有利于破坏病原菌细胞壁结构,从而发挥抑菌作用[24]。本研究通过胞外酶检测证实, 8个菌株均具有产生蛋白酶、纤维素酶、β-1,3-葡聚糖酶、木聚糖酶的能力,尤其是产蛋白酶与纤维素酶的能力较强,因而表现较好的抑制效果,推测为拮抗菌与病原菌的作用机制之一,后续可进行病原菌与拮抗菌作用机制相关研究。前人研究表明枯草芽胞杆菌YB-04、LY-1,能分泌胞外蛋白酶、淀粉酶、纤维素和β-1,3-葡聚糖酶等胞外酶,分解植物病原菌细胞壁抑制其生长[25,26],与本研究结果相符。上述研究结果证实拮抗菌通过分泌胞外酶抑制病原菌生长,为进一步明确拮抗菌与病原菌的作用机制奠定了理论基础。此外,研究发现可通过相容性试验筛选复配菌株,实现菌株之间的优势互补,增强生防效果和稳定性[27]。枯草芽胞杆菌菌株EPCO16、 EPC5与荧光假单胞菌菌株Pf1联合使用可有效抑制F. solani的生长,并提高β-1,3-葡聚糖酶、几丁质酶、酚类酶的活性,表明复配菌株间具有协同作用,增强了防治效果[28]。这一结果为探究不同菌株的协同防治效应及其在开发混合菌剂的应用研究提供理论参考。
-
![]()
图 1 烘干工艺对银耳部分营养指标的影响
注:图中不同小写字母表示同一前处理方式下不同烘干工艺之间差异显著(P<0.05),相同字母或无字母表示差异不显著(P>0.05)。
Figure 1. Effects of drying technology on nutritional indices of T. fuciformis
Note: Data with different letters indicate significant difference at 0.05 level.
表 1 不同烘干工艺下银耳的氨基酸组成
Table 1 Amino acid composition of T. fuciformis dehydrated by different drying technologies
单位:g·hg−1 氨基酸
Amino acidM1 M2 M3 M4 氨基酸
Amino acidM1 M2 M3 M4 天冬氨酸(Asp)△ 0.70±0.03 0.70±0.06 0.69±0.01 0.68±0.04 蛋氨酸(Met)* 0.19±0.02 0.18±0.02 0.19±0.01 0.17±0.03 丝氨酸(Ser)△ 0.41±0.01 0.40±0.03 0.40±0.01 0.39±0.02 赖氨酸(Lys)* 0.34±0.01 0.34±0.02 0.34±0.01 0.33±0.02 谷氨酸(Glu)△ 0.91±0.02 0.84±0.09 0.92±0.02 0.87±0.11 异亮氨酸(Ile)* 0.26±0.01 0.25±0.02 0.26±0.01 0.24±0.02 甘氨酸(Gly)△ 0.37±0.01 0.37±0.03 0.37±0.00 0.36±0.02 亮氨酸(Leu)* 0.46±0.02 0.45±0.02 0.45±0.01 0.45±0.02 丙氨酸(Ala)△ 0.40±0.02 0.40±0.02 0.40±0.01 0.38±0.02 苯丙氨酸(Phe)* 0.31±0.01 0.31±0.01 0.30±0.01 0.30±0.01 脯氨酸(Pro)△ 0.32±0.01 0.30±0.02 0.31±0.00 0.30±0.02 合计TAA 6.32±0.16 6.21±0.50 6.34±0.02 6.04±0.41 酪氨酸(Tyr) 0.28±0.03 0.33±0.05 0.32±0.04 0.29±0.04 必需氨基酸总量 TEAA 2.31±0.08 2.26±0.09 2.27±0.07 2.20±0.05 胱氨酸(Cys) 0.10±0.00 0.10±0.02 0.11±0.01 0.10±0.00 呈味氨基酸总量 TFAA 3.11±0.10 3.01±0.08 3.09±0.09 2.98±0.09 组氨酸(His)※ 0.14±0.01 0.14±0.01 0.14±0.00 0.14±0.01 TEAA/TAA(%) 36.55 37.39 35.80 36.42 精氨酸(Arg)※ 0.38±0.02 0.37±0.06 0.42±0.01 0.34±0.06 TFAA/TAA(%) 49.21 48.47 48.74 49.34 苏氨酸(Thr)* 0.39±0.01 0.38±0.02 0.38±0.01 0.37±0.02 TEAA/TNEAA(%) 57.60 58.78 55.76 57.28 缬氨酸(Val)* 0.36±0.01 0.35±0.02 0.35±0.00 0.34±0.02 注:*必需氨基酸;※儿童必需氨基酸;△呈味氨基酸;TAA氨基酸总量;TEAA必需氨基酸总量;TNEAA非必需氨基酸总量。
Note: * essential amino acid; ※ essential amino acids for children; △ flavor amino acids; TAA, total amino acid; TEAA, total essential amino acid; TNEAA, total non-essential amino acid. 下载: 导出CSV
下载: 导出CSV
表 2 烘干工艺对银耳中不同构型氨基酸组成的影响比较
Table 2 Effects of drying technology on content of amino acids of different structures in T. fuciformis
类型
TypesM1 M2 M3 M4 含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%含硫氨基酸 SAA 0.29±0.02 4.59 0.28±0.02 4.51 0.30±0.01 4.73 0.27±0.03 4.47 支链氨基酸 BCAA 1.06±0.00 16.77 1.05±0.02 16.91 1.06±0.01 16.72 1.03±0.02 17.05 芳香族氨基酸 AAA 0.59±0.01 9.34 0.64±0.01 10.30 0.62±0.01 9.78 0.59±0.02 9.77 BCAA/AAA 1.79 1.64 1.71 1.74 注:含硫氨基酸包括半胱氨酸、甲硫氨酸;支链氨基酸包括缬氨酸、异亮氨酸、亮氨酸;芳香族氨基酸包括苯丙氨酸、酪氨酸。
Note: Sulfur-containing amino acids include Cys and Met; branched chain amino acids include Val, Ile, and Leu; aromatic amino acids include Phe and Tyr.
下载: 导出CSV
表 3 烘干工艺对银耳中风味氨基酸的影响
Table 3 Effects of drying technology on amino acids of different tastes in T. fuciformis
类型
TypesM1 M2 M3 M4 含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%含量
Content/(g·hg−1)比例
Proportion/%甜味类 Sweet 1.89±0.04 29.90 1.85±0.07 29.79 1.86±0.06 29.34 1.80±0.04 29.80 苦味类 Bitter 2.10±0.05 33.23 2.05±0.07 33.01 2.11±0.05 33.28 1.98±0.06 32.78 酸味类 Sour 1.61±0.03 25.47 1.54±0.08 24.80 1.61±0.02 25.39 1.55±0.09 25.66 注:甜味类氨基酸包括甘氨酸、丙氨酸、苏氨酸、脯氨酸、丝氨酸;苦味类氨基酸包括异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、缬氨酸、组氨酸、精氨酸;酸味类氨基酸包括天冬氨酸、谷氨酸。
Note: Amino acids with sweet taste include Gly, Ala, Thr, Pro, and Ser; amino acids with bitter taste include Ile, Leu, Met, Phe, Val, His, and Arg; amino acids with sour taste include Asp and Glu.
下载: 导出CSV
表 4 不同烘干工艺制备的银耳氨基酸营养评价
Table 4 Effects of drying technology on protein nutrition of T. fuciformis
必需氨基酸 EAA 烘干方式 Drying methods 蛋 Egg FAO/WHO M1 M2 M3 M4 M1 M2 M3 M4 RAA/% RC RAA/% RC RAA/% RC RAA/% RC Ile 211 203 211 195 331 250 84.40 0.87 81.20 0.82 84.40 0.87 78.00 0.79 Leu 373 365 365 365 534 440 84.77 0.88 82.95 0.84 82.95 0.86 82.95 0.84 Lys 276 276 276 268 441 340 81.18 0.84 81.18 0.82 81.17 0.84 78.82 0.79 Met+Cys 235 227 244 219 386 220 106.81 1.44 103.18 1.05 110.90 1.15 99.54 1.01 Phe+Tyr 479 519 503 479 565 380 126.05 1.31 136.58 1.38 132.36 1.37 126.05 1.28 Thr 317 308 308 300 292 250 126.80 1.31 123.20 1.25 123.20 1.28 120.00 1.22 Val 292 284 284 276 411 310 94.19 0.98 91.61 0.93 91.61 0.95 89.03 0.90
下载: 导出CSV
表 5 不同烘干方式银耳限制性氨基酸排序
Table 5 Rank on contents of restrict amino acids in T. fuciformis
烘干工艺
Drying methods限制氨基酸 The restrict amino acid SRC 1 2 3 4 M1 Lys Ile Leu Val 80.52 M2 Lys Ile Leu Val 77.80 M3 Lys Leu Ile Val 79.15 M4 Lys Ile Leu Val 79.55
下载: 导出CSV
-
[1] ZHANG L J, WANG M S. Polyethylene glycol-based ultrasound-assisted extraction and ultrafiltration separation of polysaccharides from Tremella fuciformis (snow fungus) [J]. Food and Bioproducts Processing, 2016, 100: 464−468. DOI: 10.1016/j.fbp.2016.09.007
[2] BACH E, COSTA S, OLIVEIRA H, et al. Use of Polysaccharide extracted from Tremella fuciformis Berk for control diabetes induced in rats [J]. Emirates Journal of Food and Agriculture, 2015, 27(7): 585. DOI: 10.9755/ejfa.2015.05.307
[3] WU Y J, WEI Z X, ZHANG F M, et al. Structure, bioactivities and applications of the polysaccharides from Tremella fuciformis mushroom: A review [J]. International Journal of Biological Macromolecules, 2019, 121: 1005−1010. DOI: 10.1016/j.ijbiomac.2018.10.117
[4] 李亚欢, 田平平, 王杰, 等. 干燥方式对银耳加工与贮藏过程中品质的影响 [J]. 中国农业科学, 2016, 49(6):1163−1172. DOI: 10.3864/j.issn.0578-1752.2016.06.012 LI Y H, TIAN P P, WANG J, et al. Effects of different drying methods on quality changes during processing and storage of Tremella fuciformis [J]. Scientia Agricultura Sinica, 2016, 49(6): 1163−1172.(in Chinese) DOI: 10.3864/j.issn.0578-1752.2016.06.012
[5] 黄建立, 黄艳, 郑宝东, 等. 不同干燥方式对银耳品质的影响 [J]. 中国食品学报, 2010, 10(2):167−173. DOI: 10.3969/j.issn.1009-7848.2010.02.025 HUANG J L, HUANG Y, ZHENG B D, et al. The effect of different drying methods on the quality of Tremella fuciformis [J]. Journal of Chinese Institute of Food Science and Technology, 2010, 10(2): 167−173.(in Chinese) DOI: 10.3969/j.issn.1009-7848.2010.02.025
[6] 陈锦屏, 穆启运, 田呈瑞. 不同升温方式对烘干枣品质影响的研究 [J]. 农业工程学报, 1999, 15(3):237−240. DOI: 10.3321/j.issn:1002-6819.1999.03.049 CHEN J P, MU Q Y, TIAN C R. Study on the effect of the different heating processes on the quality of the Chinese date [J]. Transactions of the Chinese Society of Agricultural Engineering, 1999, 15(3): 237−240.(in Chinese) DOI: 10.3321/j.issn:1002-6819.1999.03.049
[7] 穆启运, 陈锦屏. 红枣挥发性物质在烘干过程中的变化研究 [J]. 农业工程学报, 2001, 17(4):99−101. DOI: 10.3321/j.issn:1002-6819.2001.04.023 MU Q Y, CHEN J P. Variation of volatile compounds of Chinese dates during toast [J]. Transactions of the Chinese Society of Agricultural Engineering, 2001, 17(4): 99−101.(in Chinese) DOI: 10.3321/j.issn:1002-6819.2001.04.023
[8] 周禹含, 毕金峰, 陈芹芹, 等. 不同干燥方式对枣粉品质的影响 [J]. 食品科学, 2014, 35(11):36−41. DOI: 10.7506/spkx1002-6630-201411008 ZHOU Y H, BI J F, CHEN Q Q, et al. Effect of drying methods on quality characteristics of jujube powder [J]. Food Science, 2014, 35(11): 36−41.(in Chinese) DOI: 10.7506/spkx1002-6630-201411008
[9] 张灵枝, 陈维信, 王登良, 等. 不同干燥方式对普洱茶香气的影响研究 [J]. 茶叶科学, 2007, 27(1):71−75. DOI: 10.3969/j.issn.1000-369X.2007.01.011 ZHANG L Z, CHEN W X, WANG D L, et al. Effect of drying methods on the aromatic character of Pu-erh tea [J]. Journal of Tea Science, 2007, 27(1): 71−75.(in Chinese) DOI: 10.3969/j.issn.1000-369X.2007.01.011
[10] 邵平, 薛力, 陈晓晓, 等. 热风真空联合干燥对银耳品质及其微观结构影响 [J]. 核农学报, 2013, 27(6):805−810. DOI: 10.11869/hnxb.2013.06.0805 SHAO P, XUE L, CHEN X X, et al. Tremella quality of combined hot-air and vacuum drying and its effect on micro-structure [J]. Journal of Nuclear Agricultural Sciences, 2013, 27(6): 805−810.(in Chinese) DOI: 10.11869/hnxb.2013.06.0805
[11] 黄艳, 郑宝东. 银耳微波真空干燥工艺优化的研究 [J]. 中国农学通报, 2009, 25(20):82−89. HUANG Y, ZHENG B D. Study on optimization of microwave vacuum drying technology for Tremella fuciformis [J]. Chinese Agricultural Science Bulletin, 2009, 25(20): 82−89.(in Chinese)
[12] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. 银耳干制技术规范: GB/T 34671—2017[S]. 北京: 中国标准出版社, 2017.. [13] 国家卫生和计划生育委员会, 国家食品药品监督管理总局. 食品安全国家标准 食品中蛋白质的测定: GB 5009.5—2016[S]. 北京: 中国标准出版社, 2017. [14] 国家卫生和计划生育委员会, 国家食品药品监督管理总局. 食品安全国家标准 食品中果糖、葡萄糖、蔗糖、麦芽糖、乳糖的测定: GB 5009.8—2016[S]. 北京: 中国标准出版社, 2017. [15] 中华人民共和国国家卫生和计划生育委员会. 食品安全国家标准 食品中还原糖的测定: GB 5009.7—2016[S]. 北京: 中国标准出版社, 2017. [16] 中华人民共和国国家卫生和计划生育委员会. 食品安全国家标准 食品中二氧化硫的测定: GB 5009.34—2016[S]. 北京: 中国标准出版社, 2017. [17] 颜孙安, 林香信, 姚清华, 等. 杂色鲍及其杂交后代的氨基酸含量和组成分析 [J]. 中国食品学报, 2013, 13(6):249−256. YAN S A, LIN X X, YAO Q H, et al. Analysis on the amino acid content and the composition of RITAI Haliotis diversicolor and their reciprocal hybrids [J]. Journal of Chinese Institute of Food Science and Technology, 2013, 13(6): 249−256.(in Chinese)
[18] 邹立扣, 潘欣, 岳爱玲, 等. 长根菇菌丝培养、鉴定及氨基酸成分分析 [J]. 食品科学, 2011, 32(3):144−147. ZOU L K, PAN X, YUE A L, et al. Cultivation, identification and amino acid composition of Xerula radicata [J]. Food Science, 2011, 32(3): 144−147.(in Chinese)
[19] 姚清华, 颜孙安, 陈国平, 等. 杏鲍菇废菌渣代料栽培对草菇营养的影响 [J]. 食品安全质量检测学报, 2019, 10(13):4314−4320. DOI: 10.3969/j.issn.2095-0381.2019.13.048 YAO Q H, YAN S A, CHEN G P, et al. Effect of spent Pleurotus eryngii substrates on qualities of Volvariella volvacea [J]. Journal of Food Safety & Quality, 2019, 10(13): 4314−4320.(in Chinese) DOI: 10.3969/j.issn.2095-0381.2019.13.048
[20] SAKLAR S, UNGAN S, KATNAS S. Microstructural changes in hazelnuts during roasting [J]. Food Research International, 2003, 36(1): 19−23. DOI: 10.1016/S0963-9969(02)00103-5
[21] 徐洲, 刘静, 冯士令, 等. 不同干燥方法对淫羊藿多糖化学性质和抗氧化活性的影响 [J]. 食品工业科技, 2015, 36(19):116−119, 123. XU Z, LIU J, FENG S L, et al. Effects of different drying methods on chemical properties and antioxidant activities of polysaccharides extracted from Epimedium [J]. Science and Technology of Food Industry, 2015, 36(19): 116−119, 123.(in Chinese)
[22] 王明, 孙曼兮, 雷激, 等. 不同干燥方式对银耳干制品品质的影响 [J]. 河南工业大学学报(自然科学版), 2015, 36(6):90−95. WANG M, SUN M X, LEI J, et al. Effects of different drying methods on the quality of dried Tremella fuciformis products [J]. Journal of Henan University of Technology (Natural Science Edition), 2015, 36(6): 90−95.(in Chinese)
[23] 丁媛媛, 毕金峰, 木泰华, 等. 不同干燥方式对甘薯产品品质的影响 [J]. 食品科学, 2011, 32(16):108−112. DING Y Y, BI J F, MU T H, et al. Effect of different drying methods on quality of sweet potato products [J]. Food Science, 2011, 32(16): 108−112.(in Chinese)
[24] 姚清华, 颜孙安, 陈美珍, 等. 古田银耳主栽品种基本营养分析与评价 [J]. 食品安全质量检测学报, 2019, 10(7):1896−1902. DOI: 10.3969/j.issn.2095-0381.2019.07.025 YAO Q H, YAN S A, CHEN M Z, et al. Basic nutrition analysis and evaluation of major cultivars of Tremella fuciformis in Gutian [J]. Journal of Food Safety & Quality, 2019, 10(7): 1896−1902.(in Chinese) DOI: 10.3969/j.issn.2095-0381.2019.07.025
[25] 顾可飞, 周昌艳. 烘干对羊肚菌营养成分影响 [J]. 食品研究与开发, 2019, 40(6):47−51. DOI: 10.3969/j.issn.1005-6521.2019.06.009 GU K F, ZHOU C Y. Effects of drying on nutritional components of morel [J]. Food Research and Development, 2019, 40(6): 47−51.(in Chinese) DOI: 10.3969/j.issn.1005-6521.2019.06.009







计量
- 文章访问数: 840
- HTML全文浏览量: 389
- PDF下载量: 25
