

Effects of Bacillus amyloliquefaciens B1619 on Growth and Fusarium Wilt of Continuous Cropping Watermelon
-
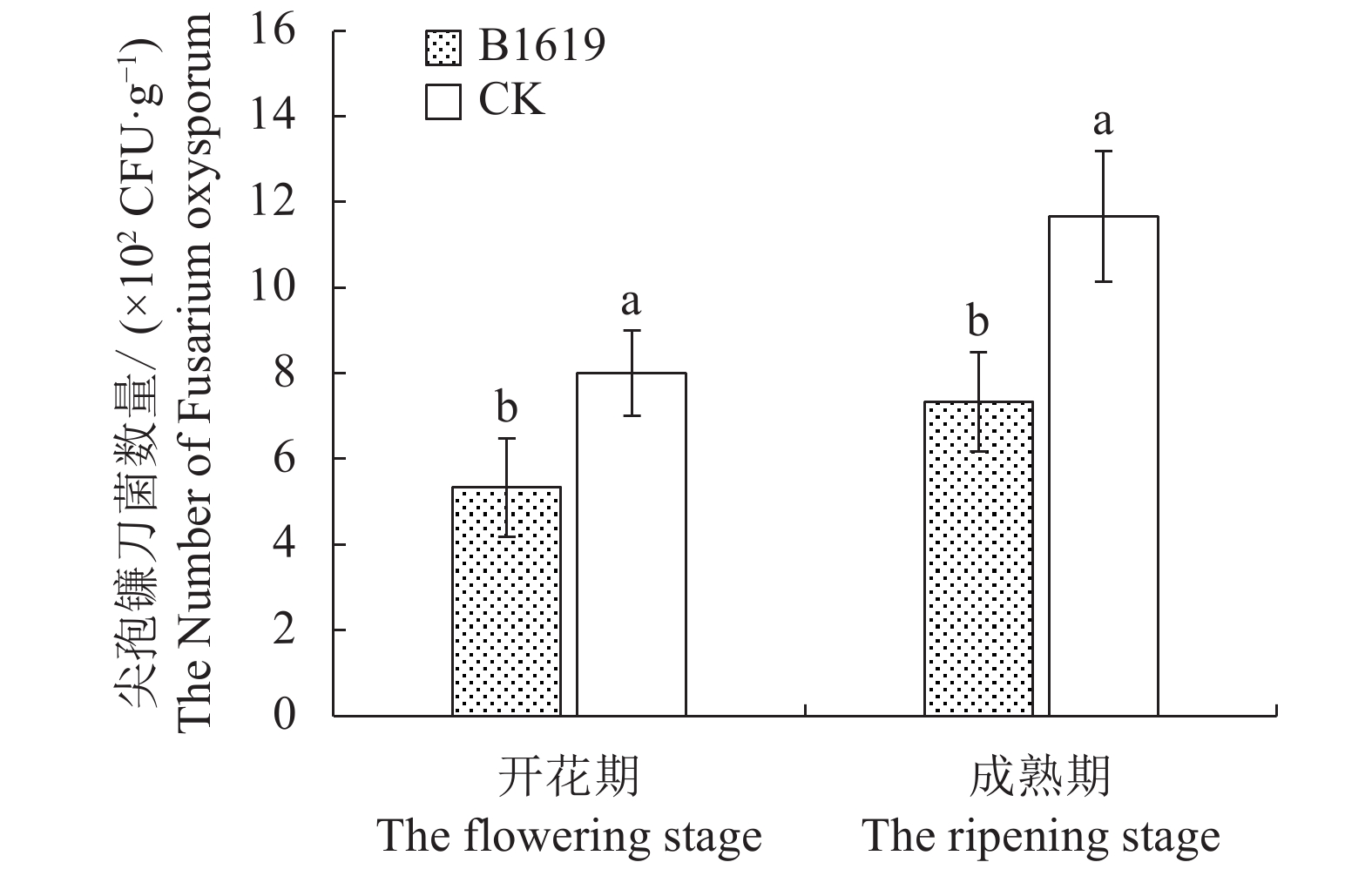
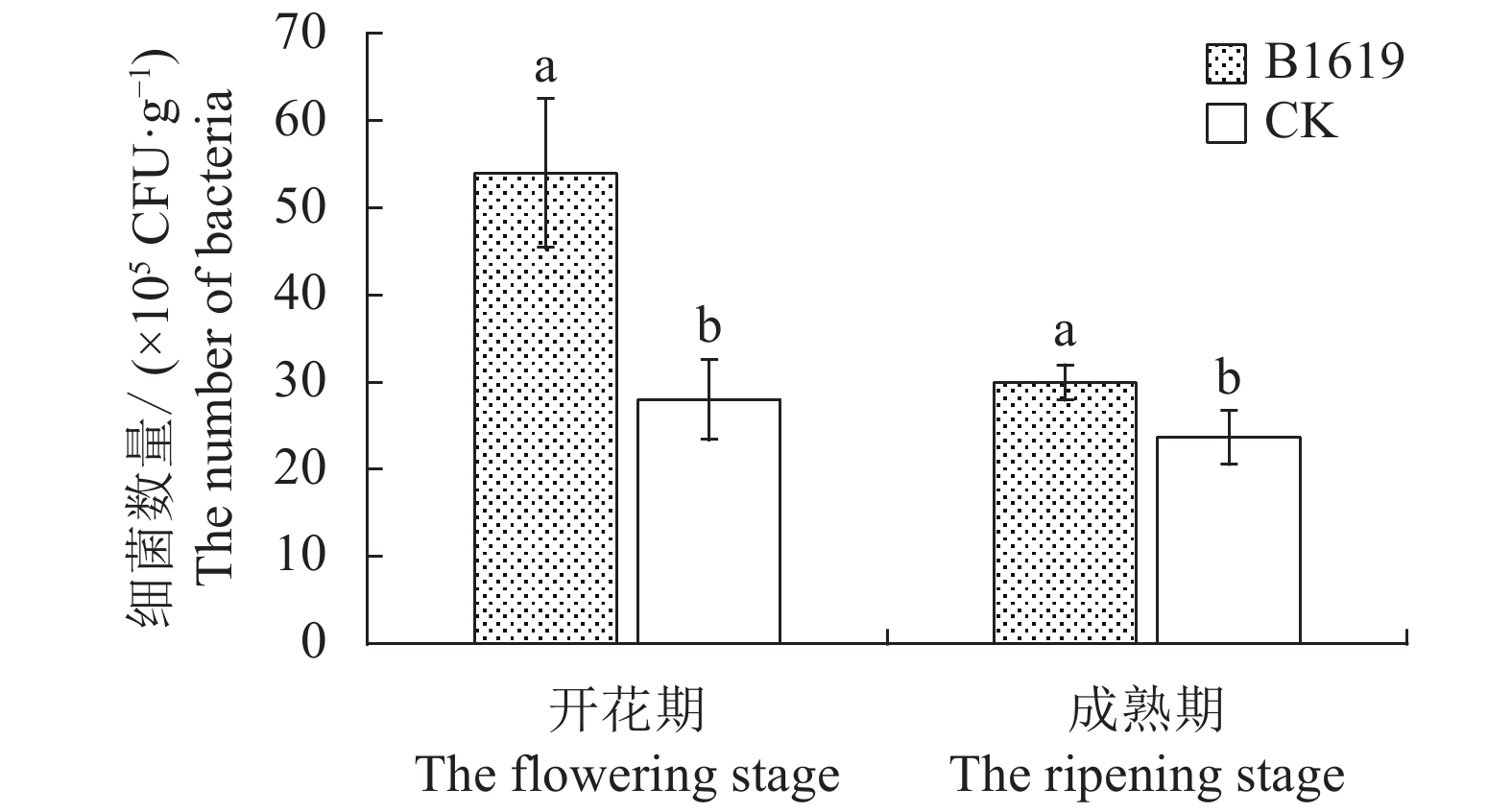
摘要:目的 明确解淀粉芽孢杆菌B1619对连作西瓜枯萎病的防效,并探讨其合理施用方法。方法 在连续种植4年西瓜的土壤环境下,采用随机区组设计,研究供试微生物菌剂在不同施用方式和施用剂量下,对连作西瓜生长、枯萎病发生及根际土壤主要菌群的影响。结果 解淀粉芽胞杆菌B1619采用穴施方式、剂量(18+9)g·穴−1时对连作西瓜的增产率达到43.20%,中心糖、边糖、商品率均显著增加。该施用方法有效改变了连作西瓜根际的菌群环境,其中细菌数量在开花期和成熟期分别是对照的1.93倍和1.27倍,病原菌尖孢镰刀菌数量分别降低为对照的33.38%和37.19%。结论 解淀粉芽胞杆菌B1619采用穴施方式、剂量(18+9)g·穴−1对4年连作西瓜产量、品质提升,西瓜枯萎病防治效果最好。Abstract:Objective Efficacy and optimum application of Bacillus amyloliquefaciens B1619 to control Fusarium wilt on continuous cropping watermelons were investigated.Methods A field experiment with a random block design was conducted for 4 consecutive years to examine the effects of the microbial agent applied by different means and dosages on the growth and disease occurrence of the watermelon and the major microflora in the rhizosphere soil.Results At a rate of (18+9) g B1619 to fill each hole dug into the ground around the plants was the selected method to deliver a 43.20% increase on watermelon yield for the study. The sugar contents in the center and peripheral by the rind of watermelon and marketability of the harvested melons increased significantly with the application. The application also changed the rhizosphere microflora community where the continuous cropping of watermelon was practiced. The bacteria count in the rhizosphere soil was 1.93 times of control at flowering stage and 1.27 times at maturing stage, while the number of pathogens, Fusarium oxysporum, reduced to 33.38% of control at flowering stage and 37.19% at maturing stage.Conclusion The “hole-digging” method of applying B1619 at (18+9) g/hole rendered a high yield, supreme fruit quality, and the much reduced incidence of Fusarium wilt on the watermelons.
-
Keywords:
- Bacillus amyloliquefaciens /
- watermelon wilt /
- application method /
- biocontrol effect
-
0. 引言
【研究意义】棕榈蓟马Thrips palmi Karny又名节瓜蓟马,属于缨翅目Thysanoptera,蓟马科Thripidae,蓟马属Thrips,分布于美国(夏威夷)、日本、印度、新加坡和中国(华南地区)等国家。主要危害茄科、葫芦科、豆科、十字花科作物,同时也是正番茄斑萎病毒属Orthotospovirus部分病毒的传播媒介[1-3]。正番茄斑萎病毒属病毒随着食物被动进入介体蓟马消化道,在消化道内建立侵染点并增殖,最终突破消化道和唾液腺屏障,病毒粒子随着唾液注入并侵染健康植株,影响植株生长、发育[4]。因此,了解消化道的组成和结构对进一步研究病毒在棕榈蓟马体内的侵染路径尤为重要。【前人研究进展】早期的缨翅目昆虫解剖学研究从形态学角度描述了缨翅目昆虫消化道、唾液腺、生殖系统等组织器官,明确不同种类蓟马的消化道形态结构存在差异[5]。此外,Ullman等利用透射电镜观察西花蓟马Frankliniella occidentalis唾液腺、前肠、中肠、后肠以及生殖系统的超薄切片,明确消化道的组织和形态结构,并探讨了蓟马消化道形态与病毒传播的关系,认为消化道形态可能决定蓟马病毒传播能力[6]。棕榈蓟马能传播番茄斑萎病毒(Tomato spotted wilt virus, TSWV)、甜瓜黄斑病毒(Melon yellow spot virus, MYSV)、花生芽枯病毒等病毒(Peanut bud necrosis virus, PBNV,)。若虫取食带毒植株5 min即可携带花生芽枯病毒,带毒成虫有效传播病毒需要取食健康植株1 h [7-10]。【本研究切入点】棕榈蓟马传播病毒的种类多,对番茄斑萎病毒、花生芽枯病毒的传播效率较高,但尚不清楚病原物在其体内的传播路径。对棕榈蓟马消化道的研究有助于更深入了解病原物的发生、复制和侵染。【拟解决的关键问题】本研究利用激光共聚焦显微镜和透射电镜在形态学和组织学上对棕榈蓟马消化道的各个部位进行研究和描述,为进一步明确病原物在消化道内的侵染路径提供依据。
1. 材料与方法
1.1 供试昆虫
棕榈蓟马于2018年4月采自云南昆明晋宁区柳坝村蔬菜大棚,鉴定后用四季豆与黄瓜饲养在人工气候箱中。饲养条件为温度27±1℃,相对湿度70%±5%,光照周期L∶D=14∶10。
1.2 消化道解剖
挑取棕榈蓟马成虫置于4℃冰箱内5~10 min,待其无活力取出放置在滴有PBS溶液(0.1 mol·L−1,pH 7.2)的载玻片上,在体视显微镜(SZ660,重庆奥特)下用镊子摘除棕榈蓟马腹部最后一节,再用镊子夹住腹部,用另外一把镊子夹住头部,将消化道从虫体中缓慢拉出,摘除多余组织。
1.3 免疫荧光样品制备
免疫荧光技术是将已知的抗原或抗体标记上荧光,再用这种荧光抗体作为探针检查细胞或组织内的相应抗原。因此,可利用该技术观察棕榈蓟马消化道组织的形态结构。将解剖出的成虫消化道迅速转移到4%多聚甲醛(Sigma)溶液中固定2 h,PBS漂洗3次后将消化道转移到2% Triton X-100(Sigma)溶液中渗透0.5 h,PBS漂洗3次后将消化道转移到含有0.5%免疫荧光染料Alexa FluorTM 633 Phalloidin(Thermo Fisher)和3%牛血清的PBS溶液中37℃孵育2 h。孵育后的消化道用PBS溶液漂洗3次,将漂洗过的消化道转移到滴有甘油的载玻片上并封片,置于Leica SP8激光共聚焦显微镜(德国Leica)下观察。
1.4 常规TEM样品制备
将解剖出的成虫消化道不同组织(前肠、中肠、后肠和马氏管)分别放入2.5%戊二醛(E-MERCK)中室温固定2 h或4℃过夜,然后用PBS溶液漂洗3次,每次15 min;再转入1%的锇酸(SPI-CHEM)溶液中固定1-2 h,PBS溶液漂洗3次,每次15 min;依次用50%、70%、80%、90%、95%的乙醇脱水,每 次15 min,最后100%乙醇和100%丙酮各脱水20 min;用spurr包埋剂(SPI-CHEM)与丙酮1∶1、1∶3分别渗透样品1 h和3 h,再用100% spurr包埋剂渗透12 h;在BJ0010聚合器(北京中兴百瑞)中70℃下聚合24 h;聚合后的样品用ULTRACUT E超薄切片机(美国AO)切片;用醋酸双氧铀(SPI-CHEM)溶液染色15 min,柠檬酸铅染色5 min,双蒸水漂洗3次,晾干后在FEI TECNAI G2透射电子显微镜(美国FEI)下观察。
2. 结果与分析
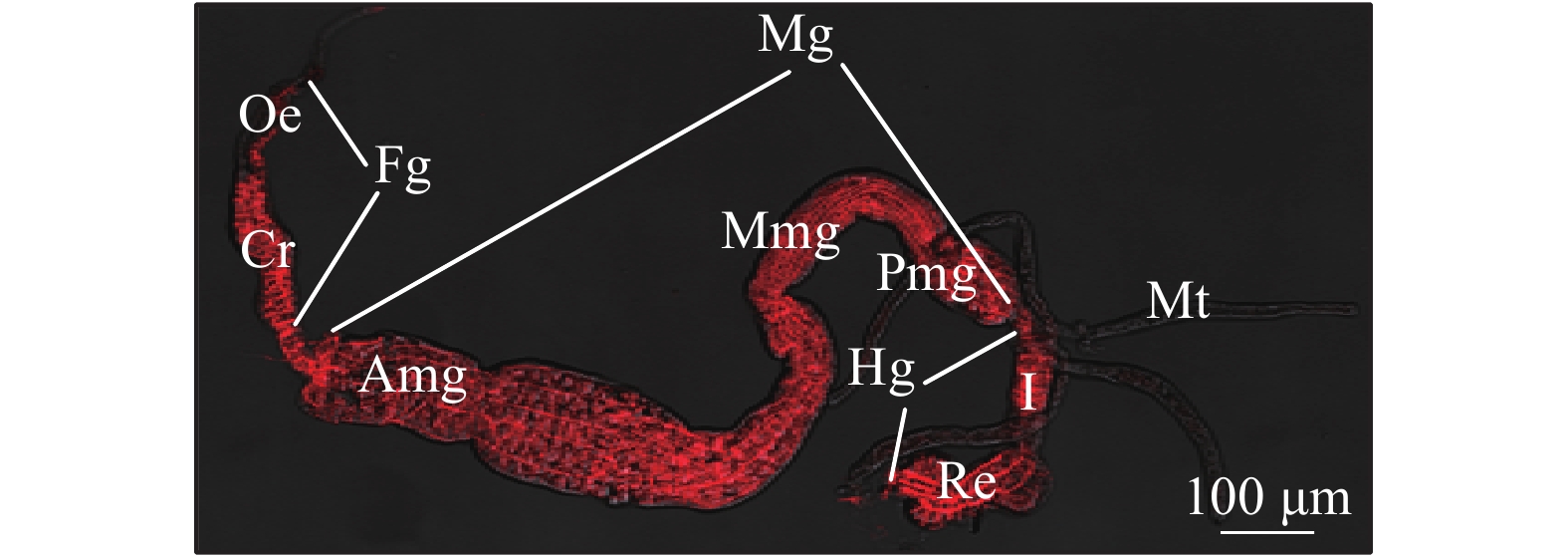
棕榈蓟马消化道始于咽部,结束于肛门,纵贯于血腔之中。根据功能不同分为前肠、中肠、后肠和马氏管(图1)。
![]() 图 1 棕榈蓟马消化道形态和结构注:Fg:前肠;Oe:食道;Cr:嗉囊;Mg:中肠;Amg:前中肠;Mmg:中中肠;Pmg:后中肠;Hg:后肠;I:回肠;Re:直肠;Mt:马氏管。Figure 1. Morphology and structure of alimentary canal of T. palmiNote: Fg: Foregut; Oe: Oesophagus; Cr: Crop; Mg: Midgut; Amg: Anterior midgut; Mmg: Middle midgut; Pmg: Posterior midgut; Hg: Hindgut; I: Ileum; Re: Rectum; Mt: Malpighian tubule.
图 1 棕榈蓟马消化道形态和结构注:Fg:前肠;Oe:食道;Cr:嗉囊;Mg:中肠;Amg:前中肠;Mmg:中中肠;Pmg:后中肠;Hg:后肠;I:回肠;Re:直肠;Mt:马氏管。Figure 1. Morphology and structure of alimentary canal of T. palmiNote: Fg: Foregut; Oe: Oesophagus; Cr: Crop; Mg: Midgut; Amg: Anterior midgut; Mmg: Middle midgut; Pmg: Posterior midgut; Hg: Hindgut; I: Ileum; Re: Rectum; Mt: Malpighian tubule.2.1 前肠(Foregut)
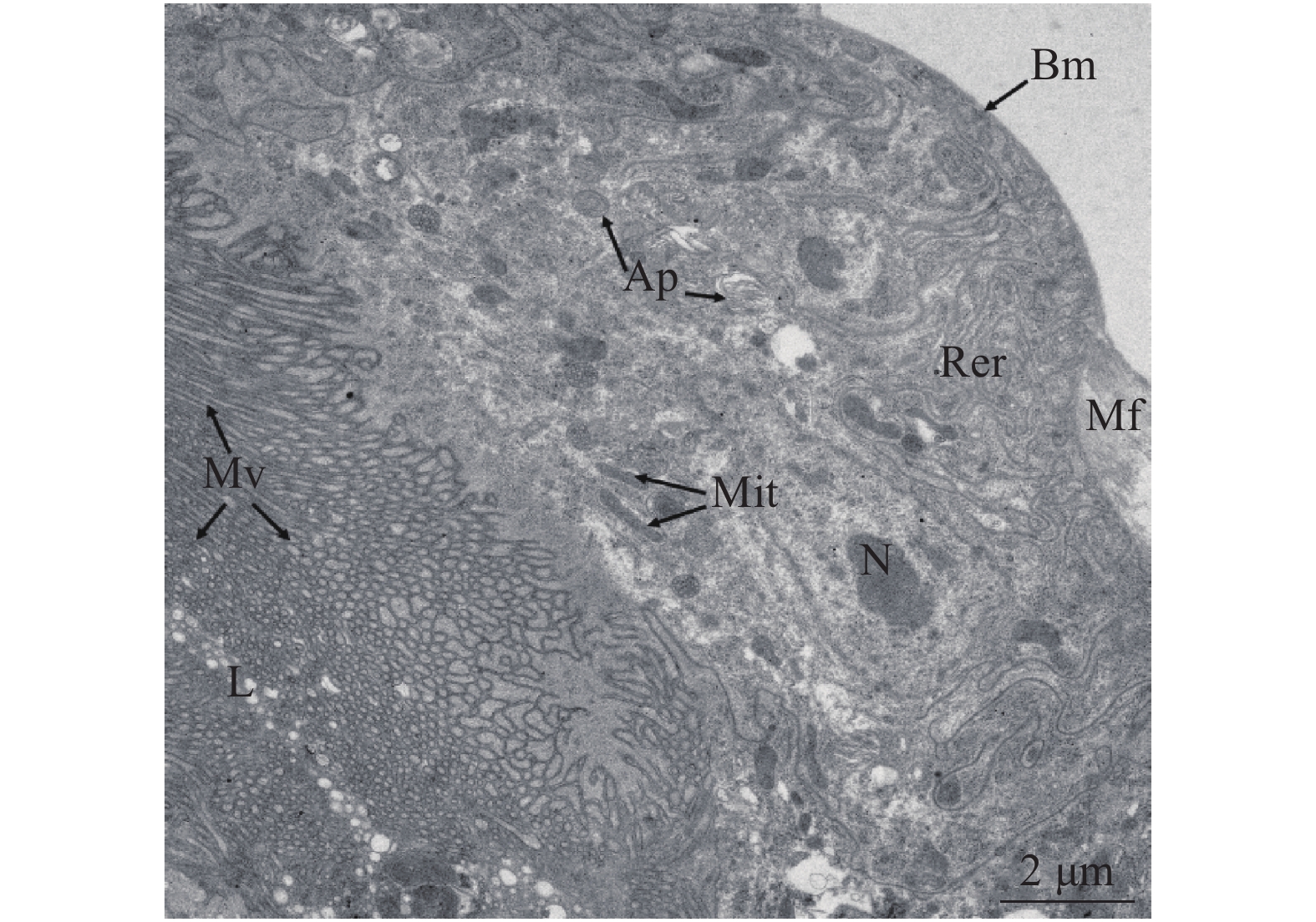
前肠位于消化道前端,长约300 ~350 μm,直径约15~25 μm,由咽、食道、嗉囊和贲门组成。食道较短、细,为膜质管状结构,肠腔由窄逐渐变宽,直到与嗉囊连接。嗉囊细胞和肠腔变大,肌肉组织丰富、膨大(图1)。嗉囊在与中肠连接处表皮凹陷,形成控制食物进出的圆环状贲门(图2)。通过透射电镜观察,发现前肠微绒毛多且密集,肠腔被微绒毛挤压呈缝隙状,线粒体分布在微绒毛附近,内质网靠近肠壁细胞的基膜(图3)。
2.2 中肠(Midgut)
中肠为棕榈蓟马消化道中最长的组织结构,长约900~1 000 μm,直径约40~120 μm,是消化食物和吸收营养的主要场所,可分为前中肠,中中肠和后中肠,以中肠在腹内的弯曲为分界线,前中肠最粗,直径约90~120 μm,其余粗细相似,直径约70~80 μm(图1)。前中肠肠壁由内向外分别为肠壁细胞、基膜、肌肉纤维,肠腔周围着生形状规则、细长且密集的微绒毛,微绒毛外包裹微绒毛膜(图4)。在微绒毛附近聚集大量的线粒体,形状为圆形或不规则的条形。细胞核形状不规则,在其周围分布大量粗面内质网(图4)。
![]() 图 2 棕榈蓟马前肠结构注:Oe:食道;Cr:嗉囊;Ca:贲门;Ec:外翻突起;Mg:中肠。图II是图I方框部位放大。Figure 2. Morphology and structure of foregut of T. palmiNote: Oe: Oesophagus; Cr: Crop; Ca: Cardia; Ec: Ectropion; Mg: Midgut. Panel II is enlarged from boxed area in Panel I.
图 2 棕榈蓟马前肠结构注:Oe:食道;Cr:嗉囊;Ca:贲门;Ec:外翻突起;Mg:中肠。图II是图I方框部位放大。Figure 2. Morphology and structure of foregut of T. palmiNote: Oe: Oesophagus; Cr: Crop; Ca: Cardia; Ec: Ectropion; Mg: Midgut. Panel II is enlarged from boxed area in Panel I.![]() 图 3 棕榈蓟马前肠超微结构注:L:肠腔;Mv:微绒毛;Ap:自噬体;Mit:线粒体;N:细胞核;Rer;粗面内质网;Bm:基底膜;Mf:肌肉纤维。Figure 3. Ultrastructure of foregut of T. palmiNote: L: Lumen; Mv: Microvilli; Ap: Autophagosome; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; Bm: Basement membrane; Mf: Muscle fiber.
图 3 棕榈蓟马前肠超微结构注:L:肠腔;Mv:微绒毛;Ap:自噬体;Mit:线粒体;N:细胞核;Rer;粗面内质网;Bm:基底膜;Mf:肌肉纤维。Figure 3. Ultrastructure of foregut of T. palmiNote: L: Lumen; Mv: Microvilli; Ap: Autophagosome; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; Bm: Basement membrane; Mf: Muscle fiber.后中肠与中中肠相似,直径较前中肠小,约40~55 μm,在末端内敛,与后肠连接(图1)。肠腔周围有微绒毛,无微绒毛膜,肠壁细胞在靠近微绒毛附近密集分布有线粒体。与前中肠不同的是肠壁褶皱状,肌肉镶嵌在褶皱中(图5)。
2.3 后肠(Hindgut)
后肠粗细居于前肠和中肠之间,长约300~400 μm,直径约30~40 μm,分为回肠和直肠,直肠连接肛门(图1)。回肠的形状为规则的管状,肠壁薄,只有一层扁平的肠壁细胞和一层肌肉纤维,内壁有一层致密的角质层,无微绒毛,角质层与细胞质膜之间有空隙,细胞内线粒体较少且分布无规则,肌肉发达,肌肉细胞与肠壁细胞厚度相似(图6)。直肠较回肠膨大,肠壁薄,细胞形态扁平,细胞核大,有类似微绒毛的长条突起,无角质层,肠腔内有大量微生物(图7)。
![]() 图 4 棕榈蓟马前中肠超微结构注:L:肠腔;Mv:微绒毛;Mvm:微绒毛膜;Mit:线粒体;N:细胞核;Rer:粗面内质网;In:基底膜内褶;Bm:基底膜;Mf:肌肉纤维。Figure 4. Ultrastructure of anterior midgut of T. palmiNote: L: Lumen; Mv: Microvilli; Mvm: Microvilli membrane; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; In: Cuticular intima lining; Bm: Basement membrane; Mf: Muscle fiber.
图 4 棕榈蓟马前中肠超微结构注:L:肠腔;Mv:微绒毛;Mvm:微绒毛膜;Mit:线粒体;N:细胞核;Rer:粗面内质网;In:基底膜内褶;Bm:基底膜;Mf:肌肉纤维。Figure 4. Ultrastructure of anterior midgut of T. palmiNote: L: Lumen; Mv: Microvilli; Mvm: Microvilli membrane; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; In: Cuticular intima lining; Bm: Basement membrane; Mf: Muscle fiber.![]() 图 5 棕榈蓟马后中肠超微结构注:①A为扁平端组织,B为褶皱端组织结构;②L:肠腔;Mv:微绒毛;Mit:线粒体;Ap:自噬体;N:细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 5. Ultrastructure of posterior midgut of T. palmiNote: ① A-Flat side of posterior midgut; B-Pleated side of posterior midgut; ② L: Lumen; Mv: Microvilli; Mit: Mitochondria; Ap: Autophagosome; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.
图 5 棕榈蓟马后中肠超微结构注:①A为扁平端组织,B为褶皱端组织结构;②L:肠腔;Mv:微绒毛;Mit:线粒体;Ap:自噬体;N:细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 5. Ultrastructure of posterior midgut of T. palmiNote: ① A-Flat side of posterior midgut; B-Pleated side of posterior midgut; ② L: Lumen; Mv: Microvilli; Mit: Mitochondria; Ap: Autophagosome; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.2.4 马氏管(Malpighian tubules)
马氏管着生于中肠与后肠的连接处,长约400~600 μm,直径约10~15 μm,共有4条,游离于血淋巴中,无分节或分支(图1)。管壁由单层的中空细胞构成,管腔周围有密集分布的特化的微绒毛,胞内细胞核和核仁都很大,紧挨微绒毛,线粒体大量分布于管壁细胞,中间有很多大小不一的囊泡,最外层为基膜,基膜上无肌肉纤维(图8)。
![]() 图 6 棕榈蓟马回肠超微结构注:Ct;角质层;Ap:自噬体;Mit:线粒体;N:细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 6. Ultrastructure of ileum of T. palmiNote: Ct: Cuticle; Ap: Autophagosome; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.
图 6 棕榈蓟马回肠超微结构注:Ct;角质层;Ap:自噬体;Mit:线粒体;N:细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 6. Ultrastructure of ileum of T. palmiNote: Ct: Cuticle; Ap: Autophagosome; Mit: Mitochondria; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.![]() 图 7 棕榈蓟马直肠超微结构注:L:肠腔;Mic:微生物;N: 细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 7. Ultrastructure of rectum of T. palmiNote: L: Lumen; Mic: Microorganism; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.
图 7 棕榈蓟马直肠超微结构注:L:肠腔;Mic:微生物;N: 细胞核;Rer:粗面内质网;Mf:肌肉纤维;Bm:基底膜。Figure 7. Ultrastructure of rectum of T. palmiNote: L: Lumen; Mic: Microorganism; N: Nucleus; Rer: Rough endoplasmic reticulum; Mf: Muscle fiber; Bm: Basement membrane.![]() 图 8 棕榈蓟马马氏管超微结构注:L:肠腔;Mv:微绒毛;V:液泡;线粒体;N:细胞核;Bm:基底膜。Figure 8. Ultrastructure of malpighian tubule of T. palmiNote: L: Lumen; Mv: Microvilli; V: Vesicles; Mit: Mitochondria; N: Nucleus; Bm: Basement membrane.
图 8 棕榈蓟马马氏管超微结构注:L:肠腔;Mv:微绒毛;V:液泡;线粒体;N:细胞核;Bm:基底膜。Figure 8. Ultrastructure of malpighian tubule of T. palmiNote: L: Lumen; Mv: Microvilli; V: Vesicles; Mit: Mitochondria; N: Nucleus; Bm: Basement membrane.3. 讨论与结论
昆虫消化道结构形态由自身的食性和发育阶段决定。不同种类的蓟马消化道在各组织的长度、宽度和形状上存在差异[5]。蓟马类昆虫以锉吸式口器吸食植物汁液,病毒首先随植物汁液进入消化道内,随后在消化道内建立侵染点并增殖,最后突破消化道和唾液腺屏障随着唾液注入并侵染健康植株[4]。本研究结果表明,棕榈蓟马消化道结构相对简单且与已报道的其他缨翅目昆虫消化道结构大体相同,主要分为前肠、中肠、后肠和马氏管,但与已报道的西花蓟马在前肠形态和中肠超微结构上有差别。棕榈蓟马前肠的食道仅起到运输食物的作用,不能进行消化和吸收,嗉囊位于前肠食道后明显变粗的部分,与食道的区别为细胞变大,肠道直径变大,尚不明确是否具有分泌消化液、初步消化食物和吸收营养物质的功能[11]。在前肠和中肠的表皮连接处有一圈突起结构,在肠腔连接处,有一环形肌肉群为贲门,在中肠蠕动时,该结构周围肌肉紧张,会使其关闭,防止食物倒流[12]。Ullman等用扫描电镜观察了西花蓟马的前肠以及周围管状结构,在对西花蓟马前肠与中肠连接处的描述中,没有观察到前肠的外翻突起,仅在贲门处有一圈向肠腔内凹陷的瓣膜,且研究中没有提到类似嗉囊的结构[6]。
中肠是昆虫消化食物和吸收营养的主要器官。本研究表明,棕榈蓟马的中肠在腹部形成两个弯曲,以增加中肠与血淋巴的营养交换面积,使营养物质交换更充分。前中肠、中中肠与后中肠的细胞形态不同,前中肠是消化道最粗的部分,细胞大,向肠腔凸起,微绒毛密集有微绒毛膜包裹,是营养吸收的主要器官。后中肠根据细胞形态可分为扁平端和褶皱端,扁平端微绒毛稀疏,肌肉处于放松状态,褶皱端紧张时微绒毛密集。在后中肠处微绒毛未发现微绒毛膜。据此推测褶皱端细胞在肌肉放松时,细胞形态与扁平端相似,在后中肠细胞中没发现Ullman等发现的同心圆环[6]。棕榈蓟马中肠肌肉比较发达,又可分为环肌和纵肌,它们垂直交叉分布,在肠道蠕动时,原本平铺在基底膜上的肌肉会横向或者纵向收缩,与其连接的细胞形态和肠道形态也会随之改变。此外,棕榈蓟马中肠无围食膜,围食膜比较发达的昆虫的食物里微生物含量比较高,因为植物汁液中微生物较少,棕榈蓟马只进化出了一层微绒毛膜[13-14]。
大部分昆虫的回肠都是由一层角质层和不发达的肠壁细胞构成,本研究表明,棕榈蓟马回肠肌肉十分发达,几乎与肠壁细胞厚度相同[15]。此外,棕榈蓟马直肠内部还有大量的微生物,这与Ullman等在西花蓟马后肠中观察到的结果一致,在电镜下的微生物形态观察也类似[6]。
不同昆虫的马氏管形态、数量都有不同,4根马氏管是缨翅目昆虫共有的特征。本研究表明,棕榈蓟马的马氏管一端与中后肠连接处链接,一端游离在血腔中。马氏管由中空的管壁细胞构成,管壁细胞内有大量的液泡,胞内的液泡被认为是由内质网或高尔基体包裹的排泄产物[16]。
综上所述,本研究利用激光共聚焦显微镜和透射电镜解剖观察了棕榈蓟马的消化道,初步明确了棕榈蓟马消化道的构成和功能,为研究病原物在棕榈蓟马体内侵染路径提供基础依据,也可以为不同种蓟马之间传毒差异比较研究提供理论支持。
-
![]()
图 1 B1619对连作西瓜根际土壤细菌数量的影响
Figure 1. Effect of B1619 on bacteria count in rhizosphere soil at continuous cropping watermelon field
![]()
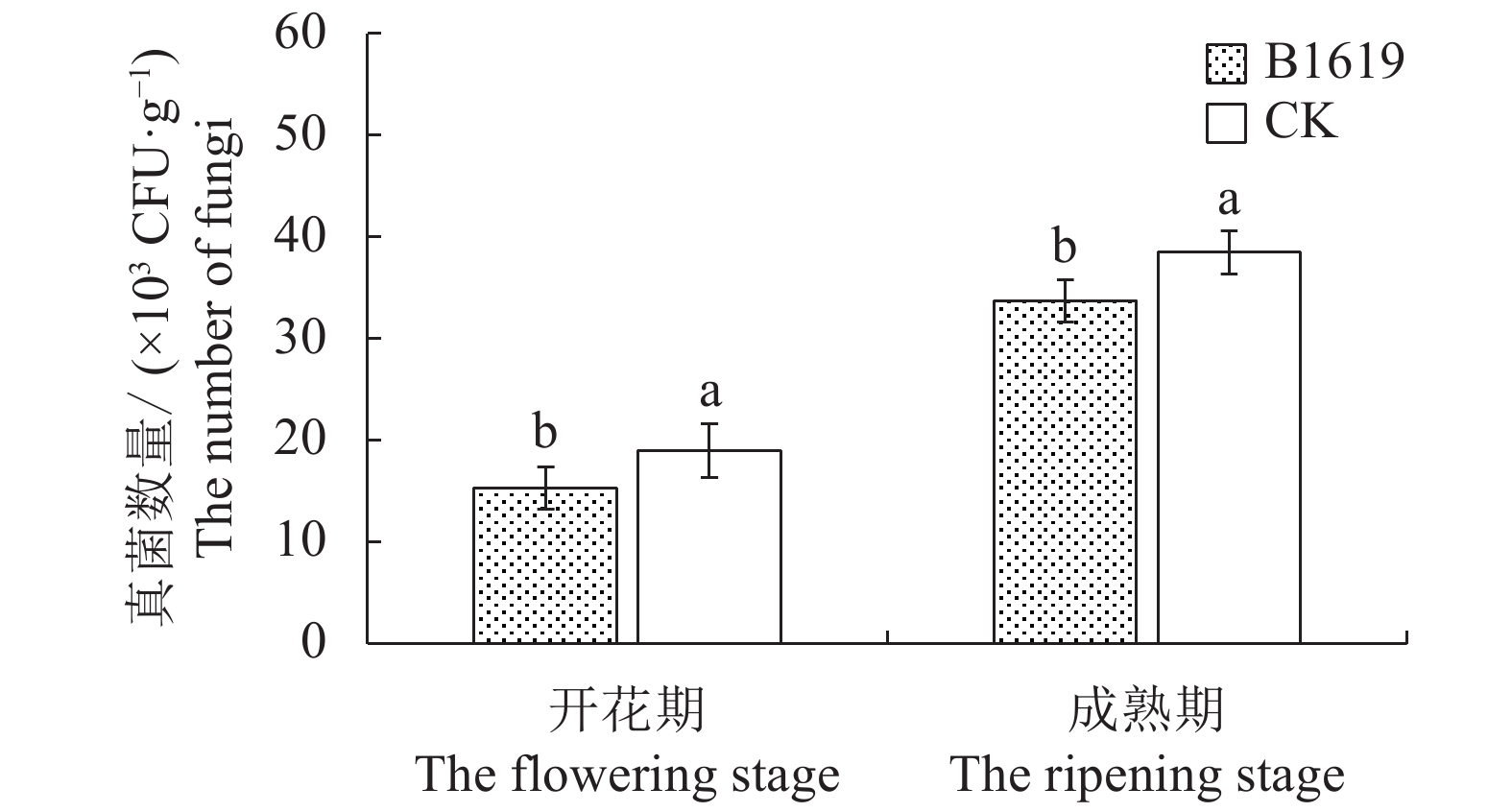
图 2 B1619对连作西瓜根际土壤真菌数量的影响
Figure 2. Effect of B1619 on fungi count in rhizosphere soil at continuous cropping watermelon field
![]()
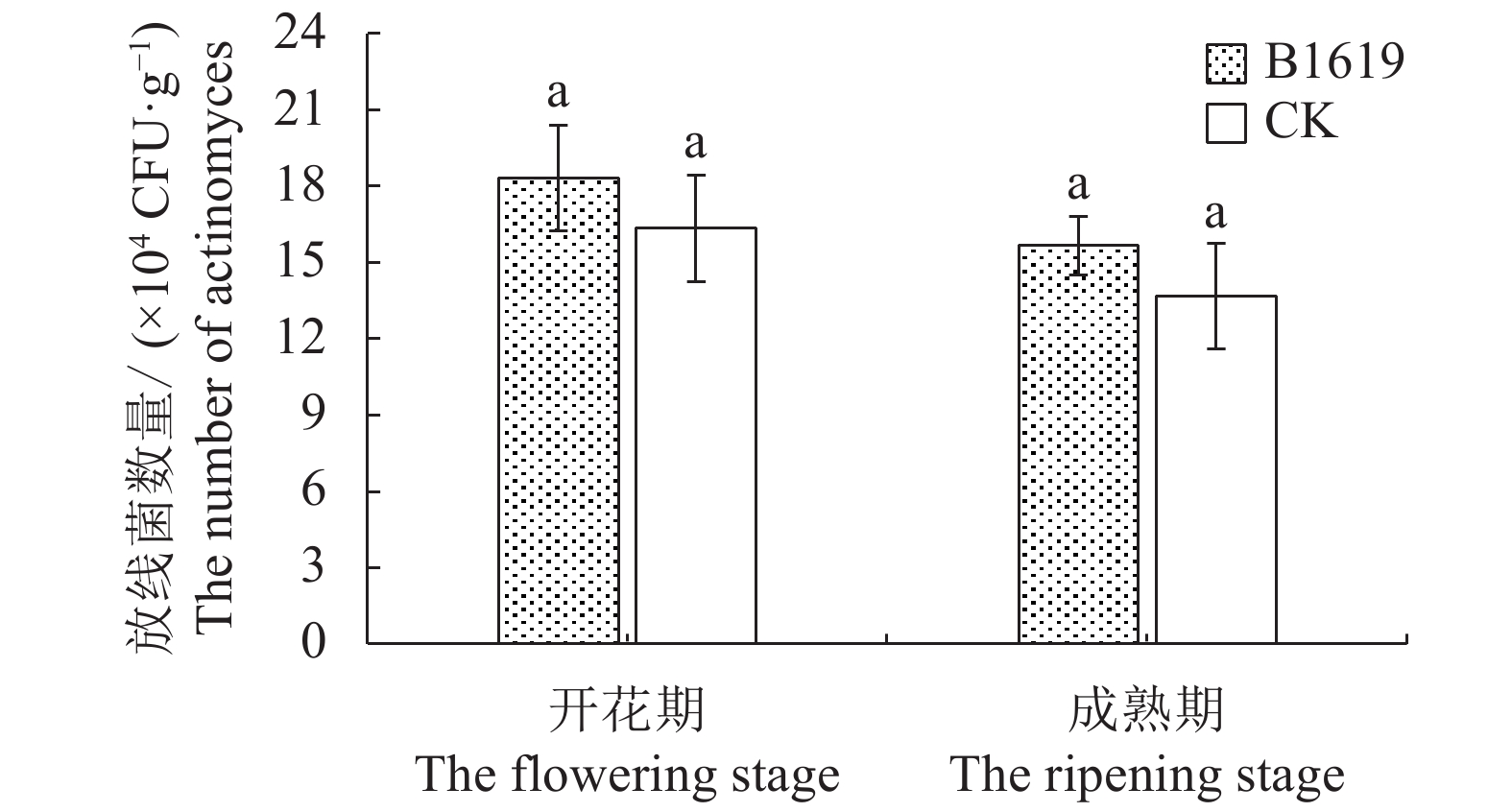
图 3 B1619对连作西瓜根际土壤放线菌数量的影响
Figure 3. Effect of B1619 on actinomyces count in rhizosphere soil at continuous cropping watermelon field
![]()
图 4 B1619对连作西瓜根际土壤尖孢镰刀菌数量的影响
Figure 4. Effect of B1619 on F. oxysporum count in rhizosphere soil at continuous cropping watermelon field
表 1 西瓜枯萎病病情分级标准
Table 1 Standards for classifying watermelon wilt
病情级数
Level of Fusarium wilt of watermelon病症
Characterization0级 植株生长正常 1级 植株萎蔫,占全株的1/4及以下,瓜果生长正常 2级 植株萎蔫,占全株的1/4至1/2,瓜皮表面稍有褪色 3级 植株萎蔫,占全株的1/2以上,茎蔓上有琥珀色胶状物,根部病茎表面产生白色或粉红色霉层,果实萎蔫缩小 4级 全株枯萎并死亡  下载: 导出CSV
下载: 导出CSV
表 2 不同施用方式和使用剂量对连作西瓜生长指标的影响
Table 2 Effect of application method and dosage of B1619 on growth of continuous cropping watermelons
处理
Treatment主蔓长
Plant height/cm茎粗
Stem diameter/mm叶面积
Leaf area/cm2叶片数
Leaf number per plant/(片·株−1)叶绿素含量SPAD值
Chlorophyll contentSPAD伸蔓期
The Stretching Stage开花期
The Flowering Stage成熟期
The Ripening Stages伸蔓期
The Stretching Stage开花期
The Flowering Stage成熟期
The Ripening Stages伸蔓期
The Stretching Stage开花期
The Flowering Stage成熟期
The Ripening Stages伸蔓期
The Stretching Stage开花期
The Flowering Stage成熟期
The Ripening Stages伸蔓期
The Stretching Stage开花期
The Flowering Stage成熟期
The Ripening StagesT1灌根
T1Root-drenching52.1 d 141.3 c 257.3 c 5.4 b 6.6 b 7.3 b 140.5 b 181.0 b 289.7 d 9.4 b 20.3 b 26.3 a 58.5 a 63.2 a 61.3 a T2灌根
T2Root-drenching67.1 b 159.8 b 304.0 a 6.2 a 7.2 a 8.0 b 203.8 a 236.3 a 370.8 b 11.8 a 22.6 a 27.9 a 56.7 a 61.5 a 58.6 a T3灌根
T3Root-drenching60.8 c 147.6 c 287.7 b 5.5 b 6.8 ab 7.7 b 151.2 b 215.8 a 339.7 c 11.0 ab 19.0 b 27.3 a 56.9 a 62.6 a 58.8 a T1穴施
T1 Hole fertilization60.1 c 144.6 c 278.3 b 5.0 b 6.6 b 7.9 b 145.2 b 214.8 a 291.6 d 10.0 b 19.2 b 28.0 a 56.9 a 62.5 a 60.0 a T2穴施
T2 Hole fertilization75.1 ab 168.0 a 309.0 a 6.6 a 7.5 a 8.8 a 233.1 a 240.7 a 390.8 b 12.6 a 19.8 b 28.3 a 55.0 a 61.9 a 57.3 a T3穴施
T3 Hole fertilization83.1 a 179.2 a 312.0 a 6.8 a 7.5 a 9.0 a 218.2 a 240.7 a 425.7 a 12.4 a 20.2 b 28.5 a 54.3 a 60.4 a 56.4 a CK 54.6 d 145.2 c 255.3 c 5.3 b 6.3 b 7.4 b 148.5 b 179.1 b 295.2 d 9.9 b 20.9 b 26.3 a 58.0 a 62.2 a 60.4 a 注: 同列数据后不同小写字母表示各处理间在0.05水平上差异显著,下同。
Note: Data followed by different lowercase letters on a same column are significantly different at 0.05 leve1. Same for the following.
下载: 导出CSV
表 3 不同施用方式和使用剂量对连作西瓜产量、品质的影响
Table 3 Effects of application method and dosage of B1619 on yield and quality of continuous cropping watermelons
处理
Treatment地上部瓜蔓鲜重
The FW of aerial part/(g·株−1)地上部瓜蔓干重
Dry weight of aerial part/(g·株−1)单瓜重
Watermelon fruit weight/kg小区产量
Plot yield/kg产量
Yield /(t·hm−2)增产率
Increase rate/%中心糖度
Central sugarcontent/%边缘糖度
Edge sugar content/%商品率
Commodity rate/%T1灌根
T1Root-drenching793.35 c 66.72 b 2.38 c 92.80 c 25.79 c 1.75 c 9.47 c 6.10 d 36.84 c T2灌根
T2Root-drenching902.05 b 70.05 b 2.70 b 107.50 b 29.88 b 17.87 b 10.84 ab 8.08 b 59.65 b T3灌根
T3Root-drenching844.60 bc 69.35 b 2.69 b 102.60 b 28.51 b 12.50 bc 10.50 b 8.20 ab 56.14 b T1穴施
T1 Hole fertilization799.50 c 66.81 b 2.57 b 98.80 bc 27.46 b 8.33 c 10.00 bc 7.13 c 54.39 b T2穴施
T2 Hole fertilization1 088.55 a 85.44 a 3.14 a 130.60 a 36.30 a 43.20 a 11.22 a 8.52 a 73.68 a T3穴施
T3 Hole fertilization1 074.20 a 78.91 a 3.23 a 133.00 a 36.96 a 45.83 a 11.18 a 8.44 a 75.44 a CK 777.36 c 59.86 c 2.37 c 91.20 c 25.35 c — 9.30 c 5.83 d 29.82 c
下载: 导出CSV
表 4 不同施用方式和使用剂量对连作西瓜病情指数的影响
Table 4 Effect of application method and dosage of B1619 on wilt control
处理
Treatment座果中期病情指数
Disease index of the fruit developing Stages成熟期病情指数
Disease index of the Ripening Stages防治效果
Disease control rate/%T1灌根
T1Root-drenching0 b 51.75 b 21.86 c T2灌根
T2Root-drenching0 b 33.33 c 49.67 b T3灌根
T3Root-drenching0 b 35.09 c 47.02 b T1穴施
T1 Hole fertilization0 b 36.84 c 44.37 b T2穴施
T2 Hole fertilization0 b 21.49 d 67.55 a T3穴施
T3 Hole fertilization0 b 21.05 d 68.21 a CK 10.07 a 66.23 a —
下载: 导出CSV
-
[1] 吴凤芝, 赵凤艳, 刘元英. 设施蔬菜连作障碍原因综合分析与防治措施 [J]. 东北农业大学学报, 2000, 31(3):241−247. DOI: 10.3969/j.issn.1005-9369.2000.03.007 WU F Z, ZHAO F Y, LIU Y Y. On the reasons of continuous cropping obstacles in vegetable facility gardening [J]. Journal of Northeast Agricultural University, 2000, 31(3): 241−247.(in Chinese) DOI: 10.3969/j.issn.1005-9369.2000.03.007
[2] 郭小敏.西瓜枯萎病菌和品种的遗传多样性研究[D].保定: 河北农业大学, 2004. GUO X M. Studies on genetic diversity with Fusarium oxysporum f.sp.niveum and cultivars[D]. Baoding: Agriculture University of Hebei, 2004.(in Chinese)
[3] 吴洪生, 周晓冬, 李鹤, 等. 黄瓜、西瓜枯萎病拮抗细菌的初步分离与鉴定 [J]. 西南农业学报, 2013, 26(3):1019−1025. DOI: 10.3969/j.issn.1001-4829.2013.03.036 WU H S, ZHOU X D, LI H, et al. Isolation and preliminary identification of antagonistic bacterium against fusarium wilt of cucumber and watermelon [J]. Southwest China Journal of Agricultural Sciences, 2013, 26(3): 1019−1025.(in Chinese) DOI: 10.3969/j.issn.1001-4829.2013.03.036
[4] WU H S, GAO Z Q, ZHOU X D, et al. Microbial dynamics and natural remediation patterns of Fusarium-infested watermelon soil under 3-yr of continuous fallow condition [J]. Soil Use and Management, 2013, 29(2): 220−229. DOI: 10.1111/sum.12029
[5] 郝文雅, 沈其荣, 冉炜,等. 西瓜和水稻根系分泌物中糖和氨基酸对西瓜枯萎病病原菌生长的影响 [J]. 南京农业大学学报, 2011, 34(3):77−82. DOI: 10.7685/j.issn.1000-2030.2011.03.014 HAO W Y, SHEN Q R, RAN W, et al. The effects of sugars and amino acids in watermelon and rice root exudates on the growth of Fusarium oxysporumf.sp.niveum [J]. Journal of Nanjing Agricultural University, 2011, 34(3): 77−82.(in Chinese) DOI: 10.7685/j.issn.1000-2030.2011.03.014
[6] KEINATH A P, HASSELL R L. Suppression of fusarium wilt caused by Fusarium oxysporum f. sp.niveum race 2 on grafted triploid watermelon [J]. Plant Disease, 2014, 98(10): 1326−1332. DOI: 10.1094/PDIS-01-14-0005-RE
[7] 沈宗专, 孙莉, 王东升,等. 石灰碳铵熏蒸与施用生物有机肥对连作黄瓜和西瓜枯萎病及生物量的影响 [J]. 应用生态学报, 2017, 28(10):3351−3359. SHEN Z Z, SUN L, WANG D S, et al. Effects of lime-ammonium bicarbonate fumigation and biofertilizer application on fusarium wilt and biomass of continuous cropping cucumber and watermelon [J]. Chinese Journal of Applied Ecology, 2017, 28(10): 3351−3359.(in Chinese)
[8] EVERTS K L, EGEL D S, LANGSTON D, et al. Chemical management of fusarium wilt of watermelon [J]. Crop Protection, 2014, 66: 114−119. DOI: 10.1016/j.cropro.2014.09.003
[9] 曹云, 宋修超, 郭德杰,等. 棉隆熏蒸与微生物有机肥联用对西瓜枯萎病的防控研究 [J]. 土壤, 2018, 50(1):93−100. CAO Y, SONG X C, GUO D J, et al. Prevention and control of watermelon Fusarium wilt by dazomet fumigation combined with bio-organic fertilizer [J]. Soils, 2018, 50(1): 93−100.(in Chinese)
[10] 吴石平, 燕嗣皇, 陆德清, 等. 木霉菌与三唑酮配合对西瓜生长的影响和对枯萎病的防效 [J]. 西南农业学报, 2002(2):65−68. DOI: 10.3969/j.issn.1001-4829.2002.02.017 WU S P, YAN S H, LU D Q, et al. Effects of triadimefon on plant growth and control of fusarium wilt in watermelon [J]. Southwest China Journal of Agricultural Sciences, 2002(2): 65−68.(in Chinese) DOI: 10.3969/j.issn.1001-4829.2002.02.017
[11] 黎起秦, 陈永宁, 林纬,等. 西瓜枯萎病生防细菌的筛选 [J]. 广西农业生物科学, 2000, 19(2):81−84. LI Q Q, CHEN Y N, LIN W, et al. Screening of bacteria strains for the biological control of watermelon wilt [J]. Journal of Guangxi Agricultural and Biological Science, 2000, 19(2): 81−84.(in Chinese)
[12] 林先贵.土壤微生物研究原理与方法[M].北京:高等教育出版, 2010. [13] 王卿. 西瓜枯萎病生防细菌的筛选及作用机制初步研究[D].南京师范大学, 2013. WAGN Q. Study on screening and mechanism of biocontrol bacteria against watermelon wilt[D]. Nanjing : Nanjing Normal University, 2013.(in Chinese)
[14] 康金磊, 韩旭, 张仕琦,等. 一株来源于茼蒿根际的西瓜枯萎病拮抗菌的鉴定及特性研究 [J]. 华北农学报, 2016, 31(S1):447−452. DOI: 10.7668/hbnxb.2016.S1.076 KANG J L, HAN X, ZHANG S Q, et al. Screening,identification and activity analysis of antagonistic bacteria against fusarium wilt of watermelon from chrysanthemum coronarium rhizosphere [J]. Acta Agriculturae Boreali-sinica, 2016, 31(S1): 447−452.(in Chinese) DOI: 10.7668/hbnxb.2016.S1.076
[15] 胡亚杰, 韦建玉, 卢健,等. 枯草芽孢杆菌在农作物生产上的应用研究进展 [J]. 作物研究, 2019, 33(2):167−172. HU Y J, WEI J Y, LU J, et al. Research progress of Bacillus subtilis application in crops production [J]. Crop Research, 2019, 33(2): 167−172.(in Chinese)
[16] 丁中, 刘跃, 徐志荣, 等. 菜丰宁B1浓缩菌粉的防病机理、生产工艺及应用技术研究 [J]. 农药, 2000, 39(8):25−27. DOI: 10.3969/j.issn.1006-0413.2000.08.011 DING Z, LIU Y, XU Z R, et al. Principle, Production and application of Bacillus subtilis(B1)for Control of Soft-rot of Chinese CabBage [J]. Pesticides, 2000, 39(8): 25−27.(in Chinese) DOI: 10.3969/j.issn.1006-0413.2000.08.011
[17] 高彦林. 微生物肥料“宁盾”大田示范推广研究[D]. 南京: 南京农业大学, 2015.. GAO Y L.Field demonstrated and research of the microbial fertilizer “Nanjing Shield”[D]. Nanjing : Nanjing Agricultural University, 2015.(in Chinese)
[18] 王璐瑶, 李兴东, 段天凤,等. 解淀粉芽胞杆菌B1619防控设施番茄枯萎病田间使用技术研究与示范 [J]. 中国生物防治学报, 2017, 33(4):512−518. WANG L Y, LI X D, DUAN T F, et al. Application technologies and demonstration trials in fields of Bacillus amyloliquefaciens B1619 against tomato fusariumWilt [J]. Chinese Journal of Biological Control, 2017, 33(4): 512−518.(in Chinese)
[19] 赵斌, 何绍江.微生物学实验[M].北京:科学出版社, 2002. [20] KOMADA H. Development of a selective medium for quantitative isolation of Fusarium Oxysporum from natural soil [J]. Review of Plant Protection Research, 1975(8): 114−125.
[21] RYU C M, FARAG M A, HU C H, et al. Bacterial volatiles promote growth in Arabidopsis [J]. Proceedings of the National Academy of Sciences, 2003, 100(8): 4927−4932. DOI: 10.1073/pnas.0730845100
[22] 杨晓云, 陈志谊, 蒋盼盼,等. 解淀粉芽孢杆菌B1619对番茄的促生作用 [J]. 中国生物防治学报, 2016, 32(3):349−356. YANG X Y, CHEN Z Y, JIANG P P, et al. Growth-promotion effect of Bacillus amyloliquefaciens B1619 on tomato plant [J]. Chinese Journal of Biological Control, 2016, 32(3): 349−356.(in Chinese)
[23] 吴勇, 黄巧云, 陈雯莉. 低温生防菌枯草芽胞杆菌BS303的选育及其效果验证 [J]. 华中农业大学学报, 2017, 36(3):19−24. WU Y, HUANG Q Y, CHEN W L, et al. Breeding and characterization of a low temperature biological control strain BS303 [J]. Journal of Huazhong Agricultural University, 2017, 36(3): 19−24.(in Chinese)
[24] 李成果. 解淀粉芽孢杆菌L3对西瓜的防病促生作用研究[D]. 南京: 南京农业大学, 2016. LI C G. The effect of Bacillus amyloliquefaciens L3 on biological control and growth-promoting of watermelon[D]. Nanjing : Nanjing Agricultural University, 2016.(in Chinese)
[25] 乔俊卿, 刘邮洲, 夏彦飞, 等. 生防菌B1619在番茄根部的定殖及对根际微生态的影响 [J]. 植物保护学报, 2013, 40(6):507−511. QIAO J Q, LIU Y Z, XIA Y F, et al. Root colonization by Bacillus amyloliquefaciens B1619 and its impact on the microbial community of tomato rhizosphere [J]. Acta Phytophylacica Sinica, 2013, 40(6): 507−511.(in Chinese)
[26] 向亚萍, 周华飞, 刘永锋, 等. 解淀粉芽孢杆菌B1619脂肽类抗生素的分离鉴定及其对番茄枯萎病菌的抑制作用 [J]. 中国农业科学, 2016, 49(15):2935−2944. DOI: 10.3864/j.issn.0578-1752.2016.15.008 XIANG Y P, ZHOU H F, LIU Y F, et al. Isolation and identification of lipopeptide antibiotics produced by Bacillus amyloliquefaciens B1619 and the inhibition of the lipopeptide antibiotics to Fusarium oxysporum f. sp. lycopersici [J]. Scientia Agricultura Sinica, 2016, 49(15): 2935−2944.(in Chinese) DOI: 10.3864/j.issn.0578-1752.2016.15.008
[27] 张斌, 杨晓云, 刘邮洲, 等. 江苏省3个番茄种植基地枯萎病菌种群数量监测及生防菌B1619的控病效果 [J]. 西南农业学报, 2015, 28(6):2521−2526. ZHANG B, YANG X Y, LIU Y Z, et al. Monitoring population dynamics of tomato fusarium wilt and control effect of bio-control bacteria B1619 at 3 tomato planting bases in jiangsu province [J]. Southwest China Journal of Agricultural Sciences, 2015, 28(6): 2521−2526.(in Chinese)
[28] BLOM D, FABBRI C, CONNOR E C, et al. Production of plant growth modulating volatiles is widespread among rhizosphere bacteria and strongly depends on culture conditions [J]. Environmental microbiology, 2011, 13(11): 3047−3058. DOI: 10.1111/j.1462-2920.2011.02582.x
-
期刊类型引用(2)
1. 王谅,刘宇艳,李恒,陈艺欣,林硕,余芸,田厚军,林涛,张洁,陈勇,魏辉. 豆大蓟马消化道形态及超微结构. 昆虫学报. 2022(02): 144-156 .  百度学术
百度学术
2. 王小奇,梁舒勍,鲁莹. 6种蝗虫消化道的超微结构与食性相关性研究. 信阳师范学院学报(自然科学版). 2020(04): 553-559 . 百度学术
其他类型引用(1)






计量
- 文章访问数: 1422
- HTML全文浏览量: 481
- PDF下载量: 23
- 被引次数: 3
