

Polymorphism of IGFBP2 Gene Relating to Meat Quality of Sujiang Pigs
-
摘要:目的 探讨IGFBP2基因潜在的遗传变异及其与苏姜猪肉质性状之间的关系。方法 采用PCR-RFLP技术检测IGFBP2基因在苏姜猪、姜曲海猪、杜洛克猪中的Msp Ⅰ酶切遗传多态性,采用单因素方差分析法分析该多态位点对苏姜猪肉质性状的影响。结果 在3个猪种试验群体,IGFBP2基因第2内含子内均发现了1个Msp Ⅰ酶切多态性,存在A、B等位基因,AA、AB、BB基因型,优势等位基因为B,优势基因型为AB,多态信息含量均呈现中度多态。苏姜猪试验群体IGFBP2基因AA、AB型个体大理石纹显著高于BB型(P<0.05),AA型个体五分制肉色和色差仪测得的a值均显著高于AB、BB型(P<0.05)。结论 苏姜猪试验群体IGFBP2基因第2内含子内PCR-RFLP-Msp Ⅰ多态性与部分肉质性状间存在一定程度的显著性相关,可以作为与猪肉质性状相关的候选基因加以研究。
-
关键词:
- 猪 /
- IGFBP2基因 /
- PCR-RFLP-Msp Ⅰ /
- 肉质性状 /
- 关联分析
Abstract:Objective Genetic variation of the insulin-like growth factor binding protein 2 gene (IGFBP2 gene), relating to the meat quality of Sujiang pigs was studied.Method The polymorphism of IGFBP2 gene was detected by PCR-RFLP in Sujiang pigs, Jiangquhai pigs and Duroc pigs, and the association between IGFBP2 gene polymorphism and meat quality traits in Sujiang pigs was analyzed.Result One common Msp Ⅰ polymorphism in the intron 2 of IGFBP2 gene was found in the 3 pig populations. It consisted of 3 genotypes (AA, AB and BB) and two alleles (A and B) with AB being the dominant genotype and B the prevailing allele. The polymorphisms of the genes from the 3 pig species were moderate on their information content. The meat marbling of Sujiang pigs with the AA and AB genotypes was significantly more than that with the BB genotypes ( P< 0.05). The meat color of the pigs with the AA genotype was significantly more intense than that with the AB and BB genotypes ( P<0.05).Conclusion The PCR-RFLP- Msp I polymorphism in the intron 2 of IGFBP2 gene of Sujiang pigs appeared to significantly correlate with the marbling and color of the pork. Consequently, the gene could potentially be used to study the meat quality improvement through breeding.-
Keywords:
- pig /
- IGFBP2 gene /
- PCR-RFLP-Msp Ⅰ /
- meat quality /
- correlation
-
0. 引言
【研究意义】山药为薯蓣科Dioscoreaceae薯蓣属Dioscorea L.多个种的统称,是一种药粮菜兼用的高效经济作物,其肉质块茎富含蛋白质、氨基酸、多糖、黏蛋白、皂苷、尿囊素等成分,具有养胃健脾、降血糖、降血脂、抗氧化、抗衰老、增强免疫和抗肿瘤等功效[1],既可鲜食、药用,又可加工成山药粉、面、酒等,利用空间广阔。福建省山药种植历史悠久,山地、平原均可种植,近年来随着需求量的上升更是得到了快速发展,成为当地脱贫致富的重要产业。【前人研究进展】福建省地方山药品种资源丰富,但鉴定和评价方面的研究较少,而资源评价与鉴定是种质资源合理开发的基础,有关福建山药种质资源的研究主要集中在农艺性状、成分分析和采用分子标记评价遗传多样性[2-6],虽有对福建山药资源的表型性状及遗传多样性报道[7],但评价指标较少,未涉及单株鲜重等产量关键指标,未见对福建山药地方品种资源进行较系统的收集和评价。【本研究切入点】调研发现2018年福建全省的山药栽培面积达5 000 hm2以上,且产值可观,但仍存在育成的新品种较少、品种更新换代慢、地方主栽品种种质退化等问题。同物异名、同名异物现象突出,特别是山药的基原多,同科不同属,其化学成分和营养品质不同,严重影响了山药的生产、收购和供应,影响了山药地方品种的利用,需加强对福建山药地方品种资源的收集与评价,开展多样性分析,明确其基原类别,筛选出高产、抗逆、外观品质优良的山药品种[8]。【拟解决的关键问题】本研究通过对福建山药地方品种资源进行收集评价,调查统计不同地方品种资源的地上部茎、叶、花、长势及地下部块茎的形态指标,采用多样性分析、相关性分析、主成分分析和聚类分析,旨在为福建山药地方品种资源的科学利用和品种选育提供科学依据。
1. 材料与方法
1.1 试验材料
2018年收集福建省南平、龙岩、三明、宁德、泉州等地区的地方主栽山药品种资源,共37份(表1)。其中包括19份褐苞薯蓣Dioscorea persimilis Prain et Burkill、3份薯蓣Dioscorea opposita Thunb.、3份山薯Dioscoreaf ordii Prain et Burkill和12份参薯Dioscorea alata L.。
表 1 供试福建山药地方资源名称、来源及基原Table 1. Names, sources and origins of Chinese yams in Fujian编号 No 俗称 General name 来源 Source 基原 Origins 1 硬壳薯 南平建阳区麻沙镇江坝村 褐苞薯蓣 2 土薯 南平建阳区麻沙镇江坝村 3 大叶永安薯 南平建阳区麻沙镇江坝村 4 小叶永安薯 南平建阳区麻沙镇江坝村 5 宣和雪薯 龙岩连城县宣和乡前进村 6 长汀淮山 龙岩长汀县古城镇南岩村 7 土薯 三明宁化县水茜乡石寮村 8 宁化早熟 三明宁化县曹坊镇三黄村 9 宁化晚熟 三明宁化县曹坊镇三黄村 10 奶薯 三明宁化县曹坊镇三黄村 11 徐引1号 三明宁化县曹坊镇三黄村 12 徐引2号 三明宁化县曹坊镇三黄村 13 清流雪薯 三明清流县林畲乡孙坊村 14 土薯 三明明溪县城关乡大坪村 15 明溪淮山1号 三明明溪县城关乡狮窠村 16 安砂小薯 三明永安市安砂镇茶仔林村 17 麻沙薯 宁德屏南县熙岭乡秀溪村 18 土薯 泉州永春县井斗镇锦溪村 19 山格淮山 泉州安溪县长坑乡山格村 20 江西薯 南平建阳区麻沙镇江坝村 山薯 21 野药薯 龙岩长汀县古城镇南岩村 22 张嘴乐淮山 宁德屏南县熙岭乡秀溪村 23 六月薯 南平建阳区麻沙镇江坝村 薯蓣 24 七月薯 南平建阳区麻沙镇江坝村 25 绵薯 宁德屏南县熙岭乡大塅村 26 黑鬼 南平建阳区麻沙镇江坝村 参薯 27 大薯 南平建阳区麻沙镇江坝村 28 紫薯 南平建阳区麻沙镇江坝村 29 红皮白肉 龙岩连城县宣和乡前进村 30 紫薯 龙岩长汀县古城镇南岩村 31 红皮白肉 龙岩长汀县古城镇南岩村 32 紫薯 三明宁化县曹坊镇三黄村 33 红皮白肉 三明明溪县城关乡大坪村 34 安砂大薯 三明永安市安砂镇培竹村 35 牛腿薯 宁德屏南县熙岭乡秀溪村 36 大薯 宁德福安市上白石镇白石板村 37 糯米薯 宁德福安市上白石镇前洋村 1.2 试验方法
1.2.1 测定指标和方法
37份山药地方品种资源按当地常规方法进行种植管理,在生长旺盛期,每份资源随机选取30株,选择离地1.5 m高的部位调查植株的生长势Growth potential(GP)、茎色Stem color(SC)、茎蔓狭翅Stem narrow-winged(SNW)、基部茎刺Stem base thorn(SBT)、叶色Leaf color(LC)、叶形Leaf shape(LS)、叶面网脉Reticulate veins on leaf surface(LSRV),测定茎粗Stem diameter(SD)、节间长Internode length(IL)、叶长Leaf length(LL)、叶宽Leaf width(LW)、叶柄长Petiole length(PL)和叶柄粗Petiole coarseness(PC);开花期观察开花与否Flowering or not(FON);采收期调查零余子有无Bulbil grow or not(BON),并随机采挖30株块茎,观察块茎形状Tuber shape(TS)、表皮光滑度Skin smoothness(SS)、须根数Number of fibrous roots(FRN)、块茎肉色Flesh color of tuber(TFC)、肉质褐变性Flesh Browning(FB),测定块茎长Tuber length(TL)、块茎直径Tuber diameter(TD)、龙头长Length of tuber top(TTL)和块茎鲜重Fresh weight of tuber(TFW),块茎直径测量选择最粗处,鲜重为采挖清洗后重量。共选取10个数量性状和14个描述型性状作为研究指标。
1.2.2 数据处理
采用Excel 2010软件进行数据整理,计算数量性状的极差、平均值、标准差、变异系数;描述型性状按标准分级赋值进行数量化(表2),计算等级分布频率,应用Shannon-Wiener index(H′)多样性指数来衡量群体多样性大小,计算公式为:
H′=−∑ii=1Pi×lnPi (Pi为某一性状第i级别内材料占总百分数的百分比,ln为自然对数)[9];应用SPSS 19.0软件进行双变量相关性分析、主成分分析和聚类分析,相关性采用Pearson相关系数表示,聚类方法采用ward’s法,度量标准采用平方Euclidean距离。表 2 表型描述性指标的分级和赋值Table 2. Grading and assignment on descriptive traits性状 Characters 分级与赋值 Grading and assignment 生长势 GP 1=弱 Weak;2=较弱 Relatively weak;3=中 Mdium;4=较强 Relatively strong;5=强 Strong 茎色 SC 1=绿 Green;2=绿带紫 Green with Purple 茎蔓狭翅 SNW 1=无 No;2=有 Yes 基部茎刺 SBT 1=无 No;2=有 Yes 叶色 LC 1=黄绿 Yellow green;2=绿 Green;3=深绿 Dark green;4=绿带紫 Green with Purple 叶形 LS 1=戟形 Halberdshape;2=长心形 Long heart shape;3=心形 Heartshape 叶面网脉 LSRV 1=不明显 Inapparent;2=明显 Apparent 零余子有无 BON 1=无 No;2=有 Yes 开花与否 FON 1=无 No;2=有 Yes 块茎形状 TS 1=短棒状 Short bar shape;2=长棒状 Long bar shape;3=短圆柱形 Short cylindrical shape;4长圆柱形 Long cylindrical shape;5=不规则块状 Irregular shape 表皮光滑度 SS 1=光滑 Smooth;2=较光滑 Relativelysmooth;3=较粗糙 Relativelyrough;4=粗糙 Rough 须根数 FRN 1=少 Only a little;2=较少 Not many;3=多 many 块茎肉色 TFC 1=白 White;2=黄白 Yellow with white;3=黄色 Yellow;4=紫色 Purple 肉质褐变性 FB 1=易 Easy;2=不易 Hard 2. 结果与分析
2.1 数量性状变异分析
对37份山药地方品种资源进行数量性状统计及变异性分析,结果见表3。由表3可知,10个数量性状的变异系数在21%~68%,由大到小依次为:块茎鲜重>龙头长>块茎长>块茎直径>茎粗>叶柄粗>节间长>叶宽>叶长>叶柄长。其中,块茎鲜重、块茎长、块茎直径的变异系数均大于30%,块茎鲜重的变异系数达68.26%。从极差看,块茎鲜重变幅最大,达2 849.14 g;其次为块茎直径和块茎长,变幅分别为75.31 mm和66.12 cm;叶长、叶宽、叶柄长和节间长的变幅均大于6 cm,说明福建山药地方品种资源块茎和叶的数量性状差异明显,地下部块茎相关性状变异范围大于地上部,遗传多样性丰富,为筛选优质资源提供了保障。
表 3 数量性状多样性统计分析Table 3. Statistical analysis on diversity of quantitative traits性状 Characters 最大值 Max 最小值 Min 极差 RI 平均值 Mean 标准差 SD 变异系数 CV/% 茎粗 SD/mm 6.30 2.09 4.21 3.19 0.89 27.90 节间长 IL/cm 14.67 6.43 8.24 10.32 2.60 25.21 叶长 LL/cm 21.18 6.63 14.55 14.53 3.44 23.66 叶宽 LW/cm 13.54 5.11 8.43 8.13 2.00 24.58 叶柄长 PL/cm 11.03 4.91 6.12 7.34 1.54 21.01 叶柄粗 PC/mm 2.87 1.16 1.71 1.72 0.44 25.67 块茎长 TL/cm 97.42 31.30 66.12 57.32 21.44 37.41 块茎直径 TD/mm 106.60 31.29 75.31 52.23 19.47 37.27 龙头长 TTL/cm 15.60 1.39 14.21 6.94 4.13 59.53 块茎鲜重 TFW/g 3 111.67 262.53 2 849.14 866.50 591.44 68.26 2.2 描述型性状遗传多样性分析
对37份山药地方品种资源的11个描述型性状进行多样性分析,类型频率分布和多样性指数见表4,由表4可知,福建山药地方品种资源描述型性状的遗传多样性指数在0.281~1.349,地上部生长势、块茎形状、表皮光滑度、须根数的多样性指数大于1。以块茎形状的多样性指数最大,基部茎刺有无最小。37份福建山药地方品种的茎蔓旋性均为右旋,各描述型性状在不同级别上分布不均匀,生长势强占半数以上,其次为中等生长势。块茎形状多样,有6种类型,以长圆柱形为主,主要为山薯、薯蓣和部分地区的褐苞薯蓣,其次为短棒状,主要为参薯。从频率分布看,福建地方品种山药多数以生长势强到中等、茎色绿、叶色绿、长心形、无零余子、有开花,块茎长圆柱形、较少须根、外表皮较粗糙、块茎肉色白、肉质不易褐变类型为主,可见福建山药地方品种资源多数生长势良好,块茎形状较齐整美观,肉色白,不易褐变,利于进一步加工保存。
表 4 描述型性状等级分布及多样性指数Table 4. Grade distribution and diversity indices on descriptive traits性状 Character 等级分布 Distribution of grade /% 多样性指数 Diversity index 1 2 3 4 5 生长势 GP 5.41 5.41 32.43 5.41 51.35 1.181 茎色 SC 75.68 24.32 0.555 茎蔓狭翅 SNW 67.57 32.43 0.630 基部茎刺 SBT 91.89 8.11 0.281 叶色 LC 5.41 75.68 16.22 2.7 0.761 叶形 LS 8.11 81.08 10.81 0.614 叶面网脉 LSRV 48.65 51.35 0.693 零余子有无 BON 75.68 24.32 0.555 开花与否 FON 40.54 59.46 0.675 块茎形状 TS 32.43 8.11 18.92 37.84 2.7 1.349 表皮光滑度 SS 54.05 18.92 27.03 1.001 须根数 FRN 10.81 21.62 48.65 18.92 1.237 块茎肉色 TFC 78.38 10.81 2.7 8.11 0.733 肉质褐变性 FB 27.03 72.97 0.584 2.3 性状相关性分析
对37份福建地方山药品种资源的24个性状进行相关性分析,其中有22个性状存在相关,结果见表5。可以看出,茎粗、叶长、叶宽、叶柄长、叶柄粗、块茎直径、块茎鲜重、生长势之间呈极显著正相关,表明叶片、叶柄长、茎粗均可以反映地上部植株生长势的强弱,地上部的生长势强弱又与地下部块茎产量密切相关,因此,可以从地上部性状的生长发育情况初步推断产量,为山药的栽培和品种选育提供依据;茎蔓具狭翅为参薯的典型特征之一,该性状与茎叶相关性状、生长势及块茎直径、块茎鲜重、块茎肉色之间呈极显著正相关,表明参薯类一般叶片更大、长势更强、产量更高、块茎肉色更丰富;叶面网脉明显与否为褐苞薯蓣的典型特征,该性状与须根数呈极显著正相关,表明褐苞薯蓣具有较多的须根;开花与否与块茎长、龙头长和块茎性状呈极显著正相关,主要是由于开花的山药类型为薯蓣、褐苞薯蓣、山薯,块茎形状以较规则的圆柱形为主,块茎长与块茎直径、龙头长相互促进,协同生长;而未观察到开花的山药主要类型为参薯,龙头较短,块茎较粗,一般呈短棒状,在土壤不够疏松条件下,更容易横向生长。
表 5 性状相关性分析Table 5. Correlation among traits项目 Items 茎粗 SD 节间长 IL 叶长 LL 叶宽 LW 叶柄长 PL 叶柄粗 PC 块茎长 TL 块茎直径 TD 龙头长 TTL 块茎鲜重 TFW 节间长 IL 0.683** 叶长 LL 0.700** 0.785** 叶宽 LW 0.819** 0.752** 0.851** 叶柄长 PL 0.565** 0.658** 0.781** 0.633** 叶柄粗 PC 0.894** 0.743** 0.854** 0.859** 0.648** 块茎长 TL −0.105 −0.061 −0.341* −0.174 −0.357* −0.25 块茎直径 TD 0.805** 0.639** 0.699** 0.710** 0.651** 0.816** −0.239 龙头长 TTL −0.464 −0.337* −0.625** −0.461** −0.566** −0.597** 0.703** −0.517** 块茎鲜重 TFW 0.770** 0.595** 0.575** 0.646** 0.487** 0.697** 0.152 0.831** −0.299 生长势 GP 0.516** 0.363* 0.588** 0.433** 0.436** 0.641** −0.365* 0.544** −0.538** 0.325* 茎蔓狭翅 SNW 0.770** 0.705** 0.875** 0.810** 0.747** 0.849** −0.447** 0.701** −0.676** 0.472** 叶色 LC −0.495 −0.135 −0.342* −0.538** −0.223 −0.455** 0.292 −0.385* 0.397* −0.298 叶形 LS 0.424** 0.25 0.573** 0.440** 0.456** 0.484** −0.444** 0.527** −0.589** 0.495** 叶面网脉 LSRV −0.738 −0.770** −0.587** −0.636** −0.447** −0.710** 0 −0.613** 0.219 −0.483** 零余子有无 BON 0.21 0.312 −0.062 −0.001 −0.094 0.128 0.385* 0.027 0.3 0.063 开花与否 FON −0.602** −0.492** −0.723** −0.657** −0.635** −0.685** 0.539** −0.648** 0.601** −0.362* 块茎形状 TS −0.138 −0.198 −0.478** −0.332* −0.363* −0.368* 0.714** −0.322 0.600** 0.138 须根数 FRN −0.504** −0.546** −0.385* −0.489** −0.509** −0.438** −0.128 −0.450** 0.03 −0.407* 表皮光滑度 SS 0.214 0.104 0.431** 0.314 0.401* 0.313 −0.690** 0.450** −0.627** 0.235 块茎肉色 TFC 0.449** 0.476** 0.524** 0.476** 0.583** 0.559** −0.334* 0.514** −0.415* 0.269 肉质褐变性 FB −0.355* −0.361* −0.19 −0.398* −0.026 −0.241 −0.211 −0.342* −0.089 −0.449** 生长势
GP茎蔓狭翅
SNW叶色
LC叶形
LS叶面网脉
LSRV零余子有无
BON开花与否
FON块茎形状
TS须根数
FRN表皮光滑度
SS块茎肉色
TFC茎蔓狭翅 SNW 0.581** 叶色 LC −0.144 −0.417* 叶形 LS 0.412* 0.489** −0.247 叶面网脉 LSRV −0.504** −0.712** 0.091 −0.064 零余子有无 BON 0.112 0.011 0.293 −0.471** −0.582** 开花与否 FON −0.494** −0.839** 0.346* −0.456** 0.518** 0.083 块茎形状 TS −0.451** −0.582** 0.252 −0.266 0.229 0.174 0.601** 须根数 FRN −0.400* −0.454** −0.022 −0.053 0.701** −0.482** 0.317 0.048 表皮光滑度 SS 0.211 0.387* −0.367* 0.652** 0.099 −0.486** −0.477** −0.521** 0.092 块茎肉色 TFC 0.385* 0.531** −0.192 0.253 −0.410* 0.025 −0.431** −0.402* −0.390* 0.334* 肉质褐变性 FB −0.159 −0.098 0.069 −0.102 0.382* −0.222 −0.007 −0.136 0.446** 0.039 −0.340* 注:**表示相关极显著(P<0.01)。
Note: **means extremely significant difference between samples (P<0.01).2.4 性状主成分分析
对24个表型性状进行主成分分析,得成分的特征值、方差和累计方差(表6)。可知第1主成分方差为43.76%,前5个成分累计方差贡献率基本达80%,说明前5个成分所包含的信息可较完整反映24个表型性状原始特征参数的大部分信息,既保留了绝大部分信息,又达到降维的目的,故提取前5个成分为筛选主要性状的依据[9]。
表 6 成分特征值及方差Table 6. Eigenvalues and variance of components成分 Component 特征值 Eigenvalues 方差 Variance/% 积累 Cumulative /% 成分 Component 特征值 Eigenvalues 方差 Variance/% 积累 Cumulative /% C1 10.501 43.755 43.755 C13 0.260 1.084 96.556 C2 4.071 16.964 60.719 C14 0.199 0.831 97.387 C3 1.878 7.825 68.544 C15 0.168 0.699 98.087 C3 1.582 6.593 75.137 C16 0.112 0.466 98.552 C5 1.080 4.502 79.639 C17 0.103 0.428 98.980 C6 0.856 3.568 83.207 C18 0.071 0.296 99.275 C7 0.740 3.084 86.292 C19 0.052 0.216 99.492 C8 0.553 2.306 88.598 C20 0.042 0.175 99.667 C9 0.526 2.191 90.788 C21 0.032 0.133 99.800 C10 0.505 2.103 92.891 C22 0.024 0.102 99.902 C11 0.316 1.318 94.209 C23 0.018 0.074 99.976 C12 0.303 1.264 95.472 C24 0.006 0.024 100.000 经方差最大正交旋转后的主成分载荷矩阵见表7。可知,第1主成分茎粗、节间长、叶长、叶宽、叶柄长、叶柄粗、块茎直径、生长势、块茎鲜重、块茎颜色、茎蔓狭翅、叶面网脉、开花与否的特征向量绝对值均大于0.6,表明第1主成分主要反应地上部茎叶长势和地下部产量构成。第2主成分中,零余子有无和块茎长的特征向量绝对值较大,分别为0.787和0.707,说明普遍具有零余子的薯蓣和山薯块茎长一般较长,同时也说明第2主成分主要反映山药的块茎长。第3主成分中,叶色特征向量绝对值最大为0.579,反映山药的叶片颜色。第4主成分中,叶形特征向量绝对值最大,为0.577,说明第4主成分主要反映山药的叶片形状。第5主成分中,块茎肉质褐变性特征向量绝对值最大,为0.558,主要反映山药肉质褐变性。各主成分的贡献率C1>C2>C3>C4>C5,即地上部长势及块茎产量构成成分>块茎长成分>叶色成分>叶形成分>块茎肉质褐变性成分。
表 7 旋转后主成分矩阵Table 7. Factor-loaded matrix after rotation特征向量 Eigen vectors C1 C2 C3 C4 C5 茎粗 SD 0.854 0.266 −0.231 0.036 0.196 节间长 IL 0.762 0.404 0.053 −0.034 0.067 叶长 LL 0.912 −0.026 0.049 0.037 0.075 叶宽 LW 0.878 0.105 −.281 −0.079 0.083 叶柄长 PL 0.791 −0.062 0.076 −0.028 −0.030 叶柄粗 PC 0.928 0.132 −0.035 0.026 0.179 块茎长 TL −0.420 0.707 −0.359 0.083 0.154 块茎直径 TD 0.866 0.113 −0.120 0.203 −0.050 龙头长 TTL −0.691 0.486 −0.241 −0.146 −0.145 块茎鲜重 TFW 0.669 0.344 −0.363 0.467 0.088 生长势 GP 0.651 0.052 0.413 0.088 −0.010 茎蔓狭翅 SNW 0.930 −0.075 0.059 −0.226 0.115 基部茎刺 SBT −0.073 0.643 0.499 0.442 −0.152 茎色 SC −0.206 0.056 0.497 0.420 0.451 叶色 LC −0.457 0.361 0.579 0.185 −0.197 叶形 LS 0.586 −0.362 −0.031 0.577 −0.085 叶面网脉 LSRV −0.697 −0.569 −0.152 0.272 −0.051 零余子有无 BON 0.005 0.787 0.299 −0.288 0.116 开花与否 FON −0.802 0.229 −0.110 0.204 −0.082 块茎形状 TS −0.506 0.506 −0.397 0.361 0.148 须根数 FRN −0.511 −0.584 −0.121 0.148 0.143 表皮光滑度 SS 0.486 −0.638 0.028 0.299 −0.218 块茎肉色 TFC 0.630 0.009 0.112 −0.135 −0.477 肉质褐变性 FB −0.286 −0.502 0.289 −0.161 0.558 2.5 性状聚类分析
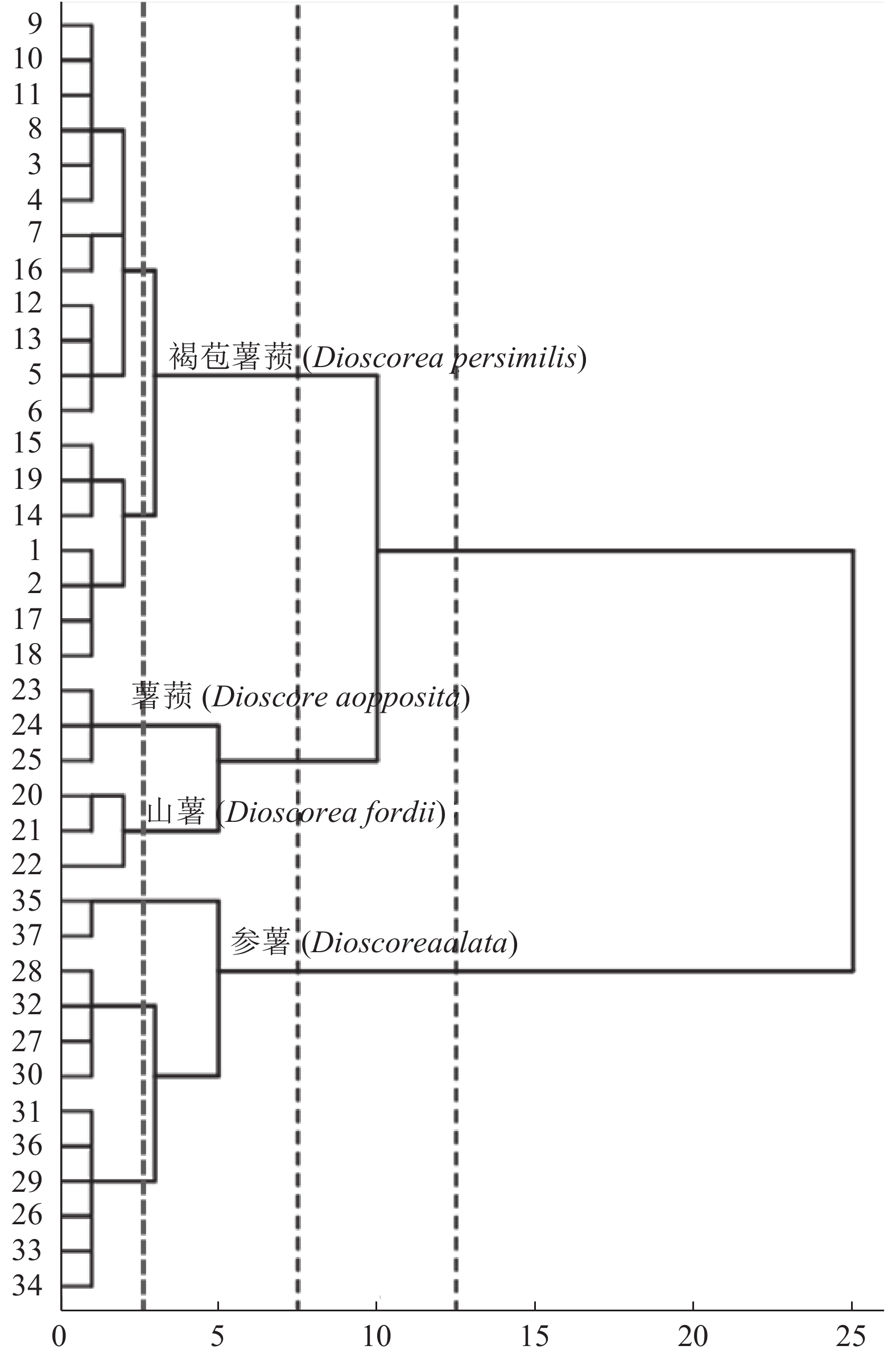
以24个性状为变量,对数据进行标准化转化,并进行聚类分析(图1)。可知,当距离为10~25时可分为2类,其中1类为参薯,该类群的遗传多样性水平最高,植株主要特征为茎蔓具狭翅、茎粗、叶长、叶宽、叶柄长、叶柄粗、生长势、块茎直径、块茎鲜重普遍较其他类别山药大;块茎肉色丰富,有白色、黄色、紫色、外紫内白等;块茎形态多样且不固定,有的在不同的土壤条件下均能垂直生长,而有些在土壤较疏松时能长成棒状,但遇到土壤较硬或干旱时,会长成脚板状、块状、牛腿状等诸多形状;仅部分资源有零余子,较少开花,在遗传距离为2.5时又可以根据块茎鲜重等继续分成3类。当距离在5~10时,可以将褐苞薯蓣从其他基原类群中分出来,该类群的主要特征为叶片中小型,叶面网脉明显,普遍开花,无零余子,地下块茎为棒状或长圆柱形,块茎茎粗较细,块茎肉色为白色,在遗传距离为2.5时又可以根据块茎鲜重、块茎直径等等继续分成2类。当距离在2.5~5.0,可以将山薯和薯蓣区分出来,山薯类群的主要特征为,叶片大小中等,叶形为狭长的心形,普遍开花,具零余子,茎蔓为绿带紫色,地下块茎为棒状或圆柱形,晚熟、抗病、产量较高。薯蓣类群的主要特征为,叶片为中小型,叶形为戟形,普遍开花、具零余子,茎蔓颜色绿带紫,早熟,地下块茎为长圆柱形,表皮较光滑。
![]() 图 1 37份山药表型性状综合聚类分析Figure 1. Comprehensive cluster analysis on phenotypic characteristics of 37 Chinese yam varieties
图 1 37份山药表型性状综合聚类分析Figure 1. Comprehensive cluster analysis on phenotypic characteristics of 37 Chinese yam varieties3. 讨论与结论
种质表型性状多样性分析能直接反应农作物商品性、产量等,因此在筛选优良株系、培育新品种中有着不可替代的作用,同时也被广泛应用于药用植物资源鉴定、遗传多样性分析和核心种质构建等,如薏苡[10]、紫苏[11]、黄连[12]等。本研究对37份福建省地方特色山药品种资源的24个表型性状进行多样性分析,可知多数资源生长势良好,块茎形状较齐整美观,刨皮后断面白且不易褐变,利于加工保存。37份山药地方品种资源的变异系数为21%~68%,其中块茎鲜重变异系数最大,多样性指数为0.281~1.349;14个描述性状中地上部生长势、块茎形状、表皮光滑度、须根数的多样性指数较大,进一步说明福建山药地方品种资源遗传多样性丰富,尤其是块茎相关性状的遗传多样性。相关性分析表明,叶片、叶柄长、茎粗均可以反映地上部植株生长势的强弱,地上部的生长势强弱又与地下部块茎产量密切相关,因此可以通过地上部长势对产量进行初步判断。
表型聚类分析将37份资源分成4个类群,即参薯类群、山薯类群、褐苞薯蓣类群、薯蓣类群,与植物分类学进行的基原分类结果一致。根据表型,薯蓣和山薯先聚为一类,再和褐苞薯蓣聚为一类,参薯的遗传距离最远,与雷伏贵等[5]、黄玉仙等[13]采用分子标记得到的亲缘关系均有差异。前人通过园艺学分类一般将山药分为普通山药和参薯,或者按块茎形状划分[14],不够严谨科学。本研究供试的37份山药资源中,褐苞薯蓣19份,参薯12份,占调查资源总数的84%,表明福建地区的褐苞薯蓣和参薯为山药的主要种植类型,与李丽红等[7]的研究基本吻合,因此参薯和褐苞薯蓣也是福建省地方药材标准收载的“福建山药”项下的基原植物[15]。但目前山薯类群在建阳、龙岩和漳州等地已有较大的种植面积[8],该基原山药也是广东省中药材标准收载的“广山药”项下的基原植物[16],为此建议将山薯类山药增收为“福建山药”的新基原类型,该类群具有晚熟,抗病性强,产量高、块茎条形顺直的优点,可从高产、不易褐变、表皮厚易运输等方面进行进一步资源筛选和推广。
-
表 1 IGFBP2基因基因型频率和基因频率
Table 1 Genotype and frequency distribution of IGFBP2 gene
品种 Breed 基因型频率 Genotype frequency 等位基因频率 Allele frequency AA AB BB A B 苏姜猪 Sujiang pig 0.16(76) 0.48(229) 0.36(172) 0.40 0.60 姜曲海猪 Jiangquhai pig 0.10(11) 0.59(64) 0.31(34) 0.39 0.61 杜洛克猪 Duroc pig 0.13(29) 0.61(132) 0.26(57) 0.44 0.56 注:括号内数字为个体数,括号外数字为频率。
Note: Number of individuals outside parentheses, frequency in parentheses. 下载: 导出CSV
下载: 导出CSV
表 2 IGFBP2基因多态位点纯合度、杂合度、有效等位基因数和多态信息含量
Table 2 Genetic polymorphism parameters ( Ho, He, Ne and PIC ) of IGFBP2 gene
品种 Breed 纯合度 H0 杂合度 He 有效等位基因数 Ne 多态信息含量 PIC 苏姜猪 Sujiang pig 0.52 0.48 1.92 0.36 姜曲海猪 Jiangquhai pig 0.52 0.48 1.92 0.41 杜洛克猪 Duroc pig 0.50 0.50 2.00 0.38 注:PIC<0.25:低度多态;0.25<PIC<0.5:中度多态;PIC>0.5:高度多态。
Note: PIC<0.25: low polymorphism; 0.25<PIC<0.5: moderate polymorphism; PIC>0.5: high polymorphism.
下载: 导出CSV
表 3 苏姜猪IGFBP2基因不同基因型肉质性状间的差异分析
Table 3 Comparison of meat quality of Sujiang pigs with varied IGFBP2 gene
基因型 Genotype pH 失水率
Water loss rate /%大理石纹评分
Marbling score肉色评分
Meat color score肉色 Meat color L a b AA 5.57±0.04 20.94±2.04 3.83±0.45a 3.87±0.23a 43.81±0.64 15.92±0.42a 20.08±0.36 AB 5.53±0.02 23.92±0.96 3.78±0.23a 2.63±0.10b 44.17±0.54 14.01±0.37b 20.39±0.27 BB 5.61±0.05 21.83±1.25 2.34±0.18b 2.73±0.15b 44.78±0.60 13.95±0.38b 20.41±0.32 注:同列数据后不同小写字母表示差异显著(P<0.05)。
Note: Different lowercase letters in the same column represented significant difference (P<0.05).
下载: 导出CSV
-
[1] SLOOTWEG M C, OHLSSON C, SALLES J P, et al. Insulin-like growth factor binding proteins-2 and-3 stimulate growth hormone receptor binding and mitogenesis in rat osteosarcoma cells [J]. Endocrinology, 1995, 136(10): 4210−4217. DOI: 10.1210/endo.136.10.7545101
[2] RAJARAM S, BAYLINK D J, MOHAN S. Insulin-like growth factor-binding proteins in serum and other biological fluids: regulation and functions [J]. Endocrine Reviews, 1997, 18(6): 801−831.
[3] ROSS M, FRANCIS G L, SZABO L, et al. Insulin-like growth factor (IGF)-binding proteins inhibit the biological activities of IGF-1 and IGF-2 but not Des-(1-3)-IGF-1 [J]. Biochemical Journal, 1989, 258(1): 267−272. DOI: 10.1042/bj2580267
[4] 杨华, 徐珍, 左波. 猪IGFBP2基因多态性及其与胴体、肉质性状的关联分析 [J]. 中国畜牧杂志, 2017, 53(3):25−28. YANG H, XU Z, ZUO B. Polymorphism of IGFBP2 Gene and Its Association with Carcass and Meat Quality Traits in Pigs [J]. Chinese Journal of Animal Science, 2017, 53(3): 25−28.(in Chinese)
[5] ROHRER G A, THALLMAN R M, SHACKELFORD S, et al. A genome scan for loci affecting pork quality in a Duroc-Landrace F2 population [J]. Animal Genetics, 2006, 37(1): 17−27. DOI: 10.1111/j.1365-2052.2005.01368.x
[6] PRASONGSOOK S, CHOI I, BATES R O, et al. Association of insulin-like growth factor binding protein 2 genotypes with growth, carcass and meat quality traits in pigs [J]. Journal of Animal Science and Technology, 2015, 57: 31. DOI: 10.1186/s40781-015-0063-3
[7] 倪黎纲, 赵旭庭, 周春宝, 等. 苏姜猪世代选育中生长、胴体和肉质性能的测定分析 [J]. 畜牧与兽医, 2015, 47(1):45−47. NI L G, ZHAO X T, ZHOU C B, et al. Determination and analysis of growth, carcass and meat quality in Sujiang pig during the generation selection [J]. Animal Husbandry & Veterinary Medicine, 2015, 47(1): 45−47.(in Chinese)
[8] 周根来, 赵旭庭, 陶勇, 等. 饲粮中消化能、粗蛋白质和粗纤维水平对育肥苏姜猪生长性能、胴体性状和肉品质的影响 [J]. 动物营养学报, 2018, 30(8):3293−3301. DOI: 10.3969/j.issn.1006-267x.2018.08.049 ZHOU G L, ZHAO X T, TAO Y, et al. Effects of dietary digestible energy, crude protein and crude fiber levels on growth performance, carcass traits and meat quality of fattening Sujiang pigs [J]. Chinese Journal of Animal Nutrition, 2018, 30(8): 3293−3301.(in Chinese) DOI: 10.3969/j.issn.1006-267x.2018.08.049
[9] 刘海霞, 朱爱文. 猪IGFBP2基因遗传多态性及其与肉质性状的关联分析 [J]. 中国畜牧兽医, 2017, 44(10):3016−3022. LIU H X, ZHU A W. Polymorphism of IGFBP2 Gene and Its Association with Meat Quality Traits in Pigs [J]. China Animal Husbandry & Veterinary Medicine, 2017, 44(10): 3016−3022.(in Chinese)
[10] 王新亮. IGFBP2通过ERK1/2信号通路调控RA-FLS细胞生物学行为机制研究[D]. 广州: 暨南大学, 2017. WANG X L. The role of IGFBP2 through ERK1/2 signal pathways regulating the RA-FLS cell biology behavior[D]. Guangzhou: Jinan University, 2017. (in Chinese)
[11] 何涛, 和姬苓. IGFBP-2结构功能与肿瘤关系研究进展 [J]. 西部医学, 2008, 20(4):858−860. DOI: 10.3969/j.issn.1672-3511.2008.04.094 HE T, HE J L. Research progress in the relationship between the structure and function of IGFBP-2 and cancer [J]. Medical Journal of West China, 2008, 20(4): 858−860.(in Chinese) DOI: 10.3969/j.issn.1672-3511.2008.04.094
[12] 许淑芬, 刘磊, 方艳秋, 等. IGFBP-2在卵巢肿瘤组织中的表达及临床意义 [J]. 中国妇幼保健, 2013, 28(27):4537−4539. DOI: 10.7620/zgfybj.j.issn.1001-4411.2013.27.40 XU S F, LIU L, FANG Y Q, et al. Expression and clinical significance of IGFBP-2 in ovarian Tumor tissue [J]. Maternal and Child Health Care of China, 2013, 28(27): 4537−4539.(in Chinese) DOI: 10.7620/zgfybj.j.issn.1001-4411.2013.27.40
[13] ZHAO X H, LI M Y, XU S S, et al. Single Nucleotide Polymorphisms in IGFBP-2 Gene and Their Associations with Body Weight Traits on Jinghai Yellow Chicken [J]. Revista Brasileira De Ciência Avícola, 2015, 17(4): 497−502. DOI: 10.1590/1516-635X1704497-502
[14] WANG W J, MENG Q Y, HU X X, et al. Chromosome location and association of haplotypes of insulin-like growth factor binding protein-2 with production performance in swine [J]. Biochemical Genetics, 2008, 46(7-8): 381−391. DOI: 10.1007/s10528-007-9128-4
-
期刊类型引用(13)
1. 马瑞婧,毕会涛,张志铭,靳珊珊,周梦丽,吴明作. 河南栾川老君山麻栎林群落结构与多样性动态研究. 河南科学. 2024(02): 242-248 .  百度学术
百度学术
2. 宋文恩,冯金露,吴文龙,申黎明,昝启杰. 大鹏半岛山油柑+水竹果+鼠刺群落多样性. 生物资源. 2024(02): 187-202 . 百度学术
3. 周建广,洪子辰,周艳,刘文芳,高亮,陈世品,林文俊. 武夷山国家公园针阔混交林群落结构与多样性研究. 武夷科学. 2024(02): 81-90 . 百度学术
4. 江子涵,高亮,周艳,郑笑,陈世品,林文俊. 武夷山国家公园甜槠林群落结构和物种多样性动态. 武夷科学. 2024(02): 91-99 . 百度学术
5. 彭泽喜,罗航,李小英,许彦红,尹五元,朱洪琴,陈玉强,陈晓琴. 滇中地区元江栲群落物种多样性及其关联因子. 林业资源管理. 2023(02): 138-146 . 百度学术
6. 孙凯,许玖玲,胡小风,厉莎,傅睿,张轩波,武舒阳. 自然保护地生态巡护道路的设计:以草海国家级自然保护区为例. 湿地科学与管理. 2023(03): 52-56 . 百度学术
7. 李洪飞,张华,伏捷,王俊夫,于立敏. 黄海北部大鹿岛鹅耳枥群落特征及其土壤环境解释. 生态学杂志. 2023(12): 2926-2935 . 百度学术
8. 李波,赵阳,齐瑞,曹家豪,陈学龙,刘婷,高本强,单立山. 海拔梯度对甘肃洮河国家级自然保护区紫果云杉林下草本植物多样性的影响. 西北林学院学报. 2022(02): 43-50 . 百度学术
9. 张伟焕,庄洁. 西藏天保工程二期生态状况动态变化分析. 林业科技通讯. 2022(05): 16-19 . 百度学术
10. 李洪飞,张华,伏捷. 黄海北部大鹿岛鹅耳枥林夏季NDVI空间分异及其环境解释. 绿色科技. 2022(13): 63-66+71 . 百度学术
11. 田莹,卢杰. 植物种群结构、数量动态与物种多样性的关系. 广西农学报. 2022(04): 78-83 . 百度学术
12. 袁刚毅,郭其强,杨瑞. 草海国家级自然保护区森林群落优势种群数量动态及物种多样性研究. 西北植物学报. 2021(07): 1229-1239 . 百度学术
13. 徐如玉,左明雪,袁银龙,孙杰,顾文杰,卢钰升,解开治,徐培智. 增施摩西管柄囊霉对甜玉米氮肥增效及土壤丛枝菌根真菌多样性的影响. 福建农业学报. 2020(04): 379-391 . 本站查看
其他类型引用(5)
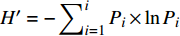

计量
- 文章访问数: 1143
- HTML全文浏览量: 377
- PDF下载量: 18
- 被引次数: 18
