

Optimization of Chlorella Farming for Shrimp Aquaculture in Aboveground Ponds
-
摘要: 介绍一种新型的高位池对虾养殖中独立培育小球藻优化模式工艺,通过单因素和正交试验设计相结合的方法研究温度、光照强度、pH、N/P(氮磷比)、Fe3+浓度等条件对小球藻定向培育的影响,并在高位池对虾养殖中进行实践应用。结果显示最佳育藻条件为温度25℃,光照7 000 lx,pH 8.5,N/P为10:1,Fe3+质量浓度500 μg·L-1,在第9~11 d小球藻有最大比生长率,可将藻液投入养殖池使用。高位池对虾养殖场的实践表明,该方法不但可以节约饲料成本,还可以较好稳定养殖水体的水质。Abstract: Chlorella farming for shrimp aquaculture in aboveground ponds was optimized by using an experiment with single factor and orthogonal design. Factors for the evaluation included light intensity and the temperature, pH, N/P ratio, and Fe3+ concentration in the water for the algae cultivation.A maximized chlorella growth coefficient was reached in 9 to 11 ds by exposing the algae to 7 000 lx light in a pH 8.5 water medium that contained 10:1 N/P nutrients and Fe3+ 500 μg·L-1 at 25℃. Thus, at that time chlorella could be harvested for feeding the shrimps. The practice could not only reduce the cost on feed for the aquaculture, but also stabilize the water quality in the shrimp pond.
-
0. 引言
【研究意义】咖啡作为云南与海南等南方省区重要的特色经济作物之一,大力发展咖啡产业对巩固脱贫攻坚成果及促进乡村振兴战略意义重大[1]。国内外咖啡传统栽培技术以覆盖耕作方式为主,即在咖啡树盘处通过覆盖秸秆、椰糠或枯枝落叶等农业废弃物,改善咖啡根际土壤微环境,提高咖啡幼苗成活率,提升咖啡产量及品质,增加经济效益[2]。然而随着咖啡种植年限的增加以及相关产业规模的扩大,传统咖啡栽培技术难于适应产业发展需求:一方面,农业覆盖材料价格的持续上涨直接提高咖啡种植成本,并且化肥农药在土壤中的累积严重威胁咖啡土壤生物健康,加剧咖啡园早衰减产现象;另一方面,咖啡初加工过程中的咖啡果皮副产物,大部分未经处理被直接丢弃,造成大量资源的浪费[3]。因此使用咖啡枯落物与果皮替代传统覆盖物,既促进咖啡枯落物与果皮的资源化利用,又减少覆盖物等投入品使用量,降低咖啡种植成本。【前人研究进展】农业废弃物作为养分的基本载体,在养分循环中是连接植物与土壤的“纽带”,在促进农田生态系统正常的物质循环和养分平衡方面有着特别重要的作用[4]。前期研究已证实咖啡枯落物与果皮等废弃物中含有丰富的氮、磷、钾元素,是制作优质有机肥的潜在材料[3],将咖啡果皮等废弃物还田分解不仅显著提升土壤养分含量,还能够改善土壤质地、提高土壤水分含量[5]。然而目前咖啡废弃物覆盖对咖啡生理机制尤其是光合特性的影响却尚不明确。一般认为作物光合作用除受光照[5, 6]、温度[7, 8]等因子影响较大外,土壤水分含量和养分含量的影响也是一个不可忽略的因素[9]。研究表明,一方面,叶片接受光照后会打开气孔,开始扩散水汽,引起叶片水势下降,降低光合作用[10]。土壤含水量的变化是影响咖啡叶片水势的主要因素[11];另一方面,矿质元素N对植物光合速率有明显的促进作用[12],并且磷和钾等元素不仅促进了叶绿素的合成,而且使叶片把所捕获的光能较充分地用于光合作用,提高了叶片的光合速率[13, 14]。但是,作物枯枝落叶等废弃物分解过程中释放的化感物质对植物叶片的光合速率等指标也具有一定的抑制作用[15],因此咖啡废弃物覆盖对咖啡叶片净光合速率、气孔导度和蒸腾速率等指标的影响目前尚未可知。此外,叶片碳利用效率(Carbon use efficiency,CUE)作为叶片碳平衡的重要指标,是光合与呼吸作用综合作用的结果,反映了咖啡碳同化能力[16],而作物水分利用效率(Water use efficiency,WUE)表征吸收单位质量的碳所消耗的水分,是衡量作物抗旱性的一个重要指标[17]。由于环境因子对咖啡叶片光合、呼吸和蒸腾的影响机制不同,环境因子与叶片碳水利用效率之间的相关关系复杂而多变[18-20],例如降水[21]、温度[22]、光照[23]和土壤养分[24]等指标均会显著改变CUE和WUE值的大小。因此,咖啡枯落物与果皮等废弃物覆盖对咖啡叶片光合指标的影响仍需进一步探究。【本研究切入点】通过明确咖啡枯落物与果皮等废弃物覆盖下土壤微环境与咖啡叶片光合指标之间的显著差异,探究不同咖啡废弃物覆盖对咖啡叶片光合能力和碳水耦合过程的其关键驱动因子。【拟解决的关键问题】探究咖啡果皮及枯落物覆盖对咖啡幼苗植株的光合速率及其碳水利用效率的影响和调控机制,为咖啡植株健康生长及其生产过程的节本增效提供理论支撑。
1. 材料与方法
1.1 试验材料
本研究采用中粒种咖啡热研1号(Coffea. Canephora Pierre ex Froehner cv. Reyan No.1)。咖啡育苗采用常规沙床催芽:将咖啡种子置于50%荫蔽度的日光温室中培育,待其长至4对真叶时,选取长势一致的植株,移栽至直径30 cm厘米的花盆中。咖啡枯落物收集于中国热带农业科学院香料饮料研究所(香饮所)咖啡种质资源圃;咖啡果皮收集于海南兴隆华侨农场太阳河咖啡加工厂(该工厂使用的咖啡品种以中粒种咖啡为主,与实验品种相同)。
1.2 试验方法
咖啡盆栽试验在2020年6月开展于香饮所种质资源圃基地(北纬18°74′,东经110°20′)。试验采用完全随机区组设计,包括未覆盖咖啡废弃物(C)、覆盖咖啡枯落物(L)、覆盖咖啡果皮(P)和同时覆盖咖啡枯落物和果皮(LP)等四种覆盖处理,每个区组内设置3个盆栽,每个处理重复6次,共72盆。作者前期随机调查海南6个咖啡种植园现存咖啡枯落物和果皮的累积量分别为301.39和239.05 g m−2。本试验采取等量添加原则,分别在L和P处理花盆中添加85.22和67.59 g咖啡枯落物和果皮,在LP处理中同时添加上述量的混合枯落物与果皮。所有盆栽咖啡苗采用相同的水肥处理,每个季度测定1次土壤与咖啡生长指标,1年后进行破坏性取样。指标测定过程如下:
生长指标:用米尺测定植株株高;叶片采集后及时测定叶面积,而后置于105 ℃杀青1 h,75 ℃烘干48 h,称重。比叶面积(SLA) = 叶干重/叶面积。光合指标的获取是在晴天上午10:00~11:00,选取茎干自下而上第4层叶,利用Li-6400XT便携式光合仪测定植株净光合速率(Pn)、叶片呼吸(R)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)等光合指标、并计算植株总光合速率(Pg)、碳利用效率(CUE)和水分利用效率(WUE),计算公式为Pg = Pn + R,CUE = Pn / Pg、WUEn = Pn / Tr、WUEg = Pg / Tr[16, 17]。每株测定3个叶片,共测定72片叶。叶绿素含量是利用SPAD-502plus型叶绿素计,在测定光合指标的同时,测定相同位叶的叶绿素相对含量(SPAD)。
1.3 数据处理
数据统计分析采用SAS V8软件进行双因素方差分析,以枯落物覆盖和咖啡果皮覆盖为两个固定因素,比较土壤理化性质指标、植株生理指标及叶片光合指标在不同处理内和处理间分别是否存在显著差异;使用CANOCO 4.5进行冗余分析(RDA),以探究影响咖啡光合指标的主要环境因子,运算过程中使用具有499个排列的蒙特卡罗测试来确定环境变量参数的显著性;Spearman等级相关性分析用于测试植株生理指标与土壤理化性质之间的关联性。运用隶属函数法对各光合和植株性状指标进行综合分析,计算出不同覆盖处理在各指标内的隶属度值,进而综合判定咖啡植株性状。隶属函数计算公式如下:
U(Xi) = (Xi−Xmin)/(Xmax−Xmin) [24]
式中:U(Xi)为隶属函数值,且U(Xi)∈[0, 1],Xi为某个光合指标测定值,Xmax、Xmin分别为该指标测定值内最大值和最小值。
使用Origin 9.8和R 4.0.3进行绘图。
2. 结果与分析
2.1 咖啡废弃物覆盖对土壤微环境及理化性质的影响
本研究中咖啡枯落物覆盖与果皮覆盖分别显著降低土壤温度0.42 ℃(F = 6.66,P<0.05)和0.33 ℃(F = 10.40,P<0.01,表1,2),但两者对土壤温度的影响不存在交互效应(表2);果皮覆盖显著提高土壤速效钾含量46.28%(F = 4.70,P<0.05),枯落物覆盖却不影响速效钾含量,且两个处理在速效钾含量的影响不存在交互效应。上述两个处理不影响土壤水分、pH、容重、有机质含量、速效磷和碱解氮含量,且在各个指标的影响不存在交互作用。
表 1 不同覆盖模式对土壤微环境及理化性质的影响Table 1. The effect of different mulching patterns on soil microclimate, physical and chemical properties种植方式 含水量
SM/%温度
ST/℃酸碱度
pH容重
SBD/N m−3有机质
SOM/g kg−1速效钾
SAK/mg kg−1速效磷
SOP/mg kg−1碱解氮
SAN/mg kg−1C 8.20±0.86 27.55±0.06 5.65±0.32 1.59±0.08 21.99±1.56 36.28±5.66 29.64±6.90 86.72±1041 L 8.33±0.88 27.33±0.05 5.57±0.30 1.61±0.10 22.10±2.77 46.42±4.78 27.74±4.81 90.84±13.97 P 7.26±0.62 27.28±0.07 5.55±0.18 1.57±0.09 21.33±3.36 33.01±7.86 28.09±8.50 91.70±17.45 P×L 8.53±1.23 27.18±0.07 5.76±0.23 1.60±0.09 21.28±2.84 54.93±10.10 33.48±9.76 90.15±13.32 注:表中数据为平均值±标准误。
Note: The data is mean ± standard errors.表 2 不同覆盖模式对土壤微环境及理化性质的双因素方差分析结果(F值)Table 2. Results of two-way ANOVA on s microclimate, physical and chemical properties under different mulching patterns (F value)种植方式 含水量
SM/%温度
ST/℃酸碱度
pH容重
SBD/N m−3有机质
SOM/g kg−1速效钾
SAK/mg kg−1速效磷
SOP/mg kg−1碱解氮SAN/mg kg−1 P 0.16 10.40** 0.07 0.08 0.00 0.13 0.05 0.01 L 0.58 6.66* 0.02 0.05 0.07 4.70* 0.07 0.02 P×L 0.38 0.82 0.31 0.00 0.00 0.64 0.22 0.04 注:表中“*”表示P<0.05, “**”表示P<0.01。
Note: “*” represent P<0.05, “**” represent P<0.01.2.2 咖啡废弃物覆盖对咖啡植株性状的影响
咖啡枯落物覆盖显著提高咖啡比叶面积45.46%(F = 6.83,P<0.05),而咖啡果皮覆盖不影响比叶面积,且两个处理对土壤温度的影响不存在交互效应(图1a);咖啡果皮覆盖显著降低咖啡株高12.11%(F = 6.39,P<0.05),枯落物覆盖却不影响株高,且两个处理对咖啡株高的影响不存在交互效应(图1c)。咖啡废弃物处理不影响咖啡叶面积指数和叶绿素相对含量,且对上述两个指标的影响不存在交互作用(图1b,d)。
![]() 图 1 不同覆盖模式对咖啡植株生理指标的影响Figure 1. Effects of different mulching patterns on physiological indexes of coffee plants
图 1 不同覆盖模式对咖啡植株生理指标的影响Figure 1. Effects of different mulching patterns on physiological indexes of coffee plants2.3 咖啡废弃物覆盖对咖啡植株光合指标的影响
咖啡枯落物覆盖与果皮覆盖不影响气孔导度、蒸腾速率、碳利用效率,却对气孔导度(F = 8.89,P<0.01)、蒸腾速率(F = 7.27,P<0.01)、碳利用效率(F = 5.19,P<0.05)的影响存在交互作用(图2a,b,f)。咖啡果皮覆盖显著提高咖啡净光合速率78.33%(F = 8.89,P<0.05)、叶片呼吸109.34%(F = 7.11,P<0.01)、总光合速率91.72%(F = 10.19,P<0.001)、净水分利用效率80.54%(F = 9.99,P<0.01)和总水分利用效率104.95%(F = 4.50,P<0.05),而咖啡枯落物覆盖不影响上述指标,且两个处理对上述指标的影响不存在交互效应(图2)。
![]() 图 2 不同覆盖模式对咖啡光合指标的影响Figure 2. Effects of different mulching patterns on photosynthetic indexes of coffee plants
图 2 不同覆盖模式对咖啡光合指标的影响Figure 2. Effects of different mulching patterns on photosynthetic indexes of coffee plants2.4 咖啡光合指标与环境因子和咖啡性状相关性分析
采用冗余分析(RDA)进一步分析各咖啡光合生理指标与环境因子之间的关系(图3)。蒙特卡洛置换检验表明(表3),土壤温度(F = 4.2,P = 0.01)对咖啡光合生理指标的影响达到显著,速效钾(F = 1.7,P = 0.06)和叶绿素相对含量(F = 1.6,P = 0.09)对咖啡光合生理指标的影响达到边际显著,所有的环境变量共同解释了样本间光合指标变异的56.7%,影响大小顺序为ST>SAK>SPAD>Height>SM>pH>SOM>SLA>LAI>SBD>SAN>SOP。RDA的前两个排序轴分别解释总方差40.26%和11.13%。
![]() 图 3 咖啡光合生理指标与环境及性状因子的冗余分析(RDA)Figure 3. Redundant analysis (RDA) of coffee photosynthetic indexes, microclimate and coffee physiological indexes表 3 环境与性状对咖啡光合生理指标的贡献及其显著性Table 3. The contributions and significances of environmental variables to soil microbial community compositions
图 3 咖啡光合生理指标与环境及性状因子的冗余分析(RDA)Figure 3. Redundant analysis (RDA) of coffee photosynthetic indexes, microclimate and coffee physiological indexes表 3 环境与性状对咖啡光合生理指标的贡献及其显著性Table 3. The contributions and significances of environmental variables to soil microbial community compositions环境与咖啡性状
Environment and coffee properties贡献率%
Contribution rate%F值 P值 土壤温度/ST 16 4.2 0.01 速效钾含量/SAK 7.3 1.7 0.06 叶绿素相对含量/SPAD 6.7 1.6 0.09 株高/Height 6.0 1.4 0.11 土壤湿度/SM 4.5 1.0 0.37 酸碱度/pH 3.6 0.8 0.45 有机质含量/SOM 2.2 0.5 0.6 比叶面积/SLA 2.4 0.5 0.62 叶面积指数/LAI 1.6 0.3 0.75 容重/SBD 1.1 0.3 0.84 碱解氮含量/SAN 1.0 0.2 0.87 速效磷含量/SOP 0.5 0.1 0.92 Spearman等级相关分析表明总光合速率(P<0.05)、净光合速率(P<0.05)、总水分利用效率(P<0.01)、净水分利用效率(P<0.05)和ST呈显著负相关;蒸腾速率、总光合速率与土壤速效钾含量之间存在显著正相关关系(P<0.05);气孔导度与叶片相对叶绿素含量之间存在显著负相关关系(P<0.01);由此可知土壤温度和速效钾等养分含量是调控咖啡光合速率等生理指标的主要环境因素(图4)。
![]() 图 4 不同咖啡废弃物覆盖下咖啡光合指标与环境因子和植株性状相关性分析注:图中“^”表示P < 0.1, “*”表示P < 0.05, “**”表示P < 0.01。Figure 4. Correlation analysis of coffee photosynthetic indexes, microclimate and plant traits indexes under different mulching patternsNote: “^” represent P < 0.1, “*” represent P < 0.05, “**” represent P < 0.01.
图 4 不同咖啡废弃物覆盖下咖啡光合指标与环境因子和植株性状相关性分析注:图中“^”表示P < 0.1, “*”表示P < 0.05, “**”表示P < 0.01。Figure 4. Correlation analysis of coffee photosynthetic indexes, microclimate and plant traits indexes under different mulching patternsNote: “^” represent P < 0.1, “*” represent P < 0.05, “**” represent P < 0.01.2.5 咖啡废弃物覆盖对咖啡光合生理指标影响综合评价
计算出各覆盖处理组在各指标内的隶属度值,进而综合判定咖啡植株生理状态的优劣。由表4可知,不同覆盖处理的平均隶属值排序从大到小为P>LP>L>C,排名最高的是咖啡果皮覆盖处理,未覆盖咖啡废弃物处理得分最低(表4)。
表 4 咖啡废弃物覆盖对咖啡光合生理指标影响综合评价Table 4. Comprehensive evaluation of effects coffee waste cover on coffee photosynthetic indexes处理 Treatment 株高 Height 叶面积指数 LAI 比叶面积 SLA 叶绿素含量 SPAD 气孔导度 Gs 蒸腾速率 Tr 净光合速率 Pn 呼吸速率 R C 0.59 0.00 0.07 1.00 0.00 0.00 0.00 0.00 L 0.00 1.00 0.00 0.11 1.00 1.00 0.77 0.10 P 1.00 0.24 0.56 0.32 0.90 0.63 1.00 1.00 LP 0.26 0.13 1.00 0.00 0.19 0.11 0.88 0.68 处理
Treatment总光合速率
Pg碳利用效率
CUE净水分利用效率
WUEn总水分利用效率
WUEg隶属度平均值
Average membership排名
RankingC 0.00 0.00 0.00 0.00 0.14 4 L 0.49 1.00 0.62 0.25 0.53 3 P 1.00 0.57 1.00 0.85 0.76 1 LP 0.80 0.62 0.95 1.00 0.55 2 3. 讨论
3.1 咖啡废弃物覆盖对土壤微环境的影响
农业废弃物作为养分的基本载体,在促进农田生态系统物质循环和养分平衡方面,维持土壤肥力方面具有重要作用[25, 26]。前期大量研究已证实土壤氮、磷和钾等养分含量随凋落物累积量的增加而显著提高[27, 28],但是本研究结果表明咖啡果皮与枯落物覆盖对土壤速效氮和磷的影响较小,与前期研究结果不一致,可能与咖啡废弃物质量有关[29]。农业废弃物质量包括易分解成分(如氮和磷化合物)和难分解成分(如木质素、纤维素和多酚类物质等)[30],在长达一年的分解过程中,氮和磷等化合物在前期短暂快速分解阶段释放,导致后期难分解成分在长期裂解过程中释放氮和磷元素不足[31]。值得注意的是,咖啡果皮覆盖显著提高土壤速效钾含量,可能与咖啡果皮中钾元素含量显著高于植株其他器官有关[5]。此外,前期的凋落物倍增实验均证实凋落物覆盖能够显著提高土壤含水量并降低土壤温度[32]。本研究的土壤温度结果与前期研究保持一致,即两种咖啡废弃物覆盖显著降低土壤温度。然而,咖啡土壤水分却不响应于废弃物覆盖处理,可能与开展盆栽试验温室的高空气湿度有关。有研究表明,农田生态系统水分散失方式主要是植物蒸腾作用[33],由于空气湿度与叶片水势之间存在正相关关系[34],温室中较高的空气湿度可能是引起咖啡蒸腾速率不响应于咖啡废弃物覆盖处理的原因之一[35, 36]。因此,在本实验的持续高土壤水分条件下,咖啡废弃物的保水功能被弱化。
3.2 咖啡废弃物覆盖对咖啡光合速率的影响
前期研究发现影响咖啡光合速率的主要因子是土壤养分、光照强度和土壤温湿度等环境条件[37-39]。本研究表明咖啡枯落物覆盖处理下光合速率的变化主要受土壤温度和土壤钾含量的调控。一方面,咖啡适宜的生长温度为22.0-28.4 ℃,同时咖啡属于浅根系植物,其对土壤温度的变化较为敏感[40]。有研究指出当土壤温度高于植物适宜光合温度,会通过抑制植株根系或叶片酶活性,进而抑制其光合速率[41-43],因此本研究中咖啡果皮覆盖能够通过降低土壤温度进而促进光合速率。另一方面,叶片钾元素能够通过改善叶肉细胞光合活性,提高净光合速率[44, 45]。咖啡果皮覆盖通过提高土壤钾的供应,进而促进叶片总光合速率的提升。
3.3 咖啡废弃物覆盖对咖啡碳水分利用效率的影响
植物的水分利用效率WUE反映的是植物由于蒸腾消耗的单位水量所能产生的光合产量。前期研究表明凋落物覆盖降低热带木本植物叶片WUE,是由于温度升高会导致植物的蒸腾速率加快从而使植物WUE降低[17, 46],导致植物WUE与温度呈显著负相关关系。而本研究结果与上述研究并不一致,原因在于咖啡果皮覆盖通过降低土壤温度促进光合速率的提升,而咖啡蒸腾速率对咖啡废弃物覆盖处理并不敏感,导致咖啡果皮覆盖下咖啡叶片在维持蒸腾速率稳定的基础上显著促进碳吸收速率,进而提高叶片WUE。本研究中土壤速效钾含量与蒸腾速率之间存在显著正相关关系,表明钾元素是加速咖啡植株水汽通量的主要驱动因子之一[47]。尽管咖啡果皮覆盖显著提高土壤速效钾含量,但咖啡蒸腾速率可能收到其他因素的干扰,其具体调控机制仍需进一步研究。
温度作为影响树木CUE最重要的气候因子,已得到广泛的关注[21, 48, 49]。前期有研究认为光合和呼吸随温度变化具有不同的敏感性,从而导致温度变化对CUE的影响存在显著差异[50]。但在本研究中碳利用效率并不响应于土壤温度的变化是由于咖啡果皮覆盖通过降低土壤温度显著提高净光合速率,但同时也显著促进呼吸速率的提高,因此对叶片瞬时碳利用效率并没有影响。上述结果可能与咖啡对温度长期变化的自适应有关[51, 52],即咖啡叶片随温度的变化,光合速率和呼吸速率会保持一个相对恒定的比值,从而恢复光合与呼吸两者之间的平衡[53]。因此不同咖啡废弃物覆盖均未改变咖啡叶片碳利用效率。
3.4 对不同废弃物覆盖下咖啡光合生理的综合性评价
单个光合指标只反映咖啡植株光合特性的某个侧面,而咖啡植株各部分的协调和平衡共同决定了咖啡的光合作用过程。本研究根据不同光合与生理指标的隶属度值,对植株的生长状态进行综合评价,其数值越高表明咖啡光合与长势越好。结果显示,单独覆盖咖啡果皮处理中咖啡植株光合指标数值最高,其与对应的株高等生长指标相符合,即为最适的咖啡废弃物覆盖处理。然而本研究中咖啡废弃物的使用量固定不变,并未考察不同量的咖啡废弃物覆盖对咖啡植株生理过程的影响。在未来的研究中,应探讨不同重量咖啡果皮与枯落物对咖啡光合指标的覆盖组合,用于获得更加科学的覆盖方案。
4. 结论
咖啡果皮覆盖通过降低土壤温度以及提高土壤速效钾含量显著提高咖啡净光合速率、叶片呼吸、总光合速率、净水分利用效率和总水分利用效率,而咖啡枯落物覆盖却不影响光合指标。经综合评价,仅使用咖啡果皮覆盖对咖啡光合指标的促进效果优于其他覆盖措施。在进行咖啡栽培实践中,使用果皮废弃物覆盖最有助于促进咖啡植株健康生长以及咖啡生产的节本增效。
-
![]()
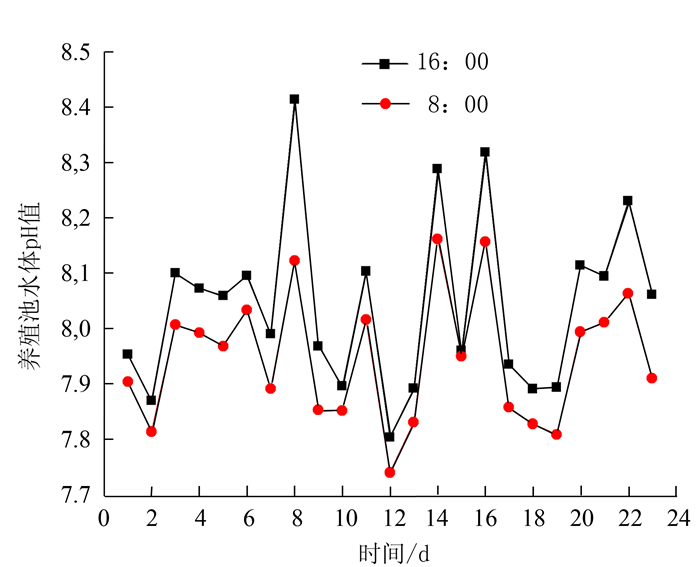
图 6 南美白对虾养殖池水体pH变化过程
Figure 6. Changes on water pH in a Penaeus vannamei aquaculture pond
表 1 正交试验因素和水平
Table 1 Factors and levels of orthogonal experiment
水平 温度/℃ 光照/lx pH N/P 铁离子/(μg·L-1) 1 15 3000 6.5 10:1 300 2 20 4000 7.0 20:1 400 3 25 5000 7.5 30:1 500 4 30 6000 8.0 40:1 600 5 35 7000 8.5 50:1 700  下载: 导出CSV
下载: 导出CSV
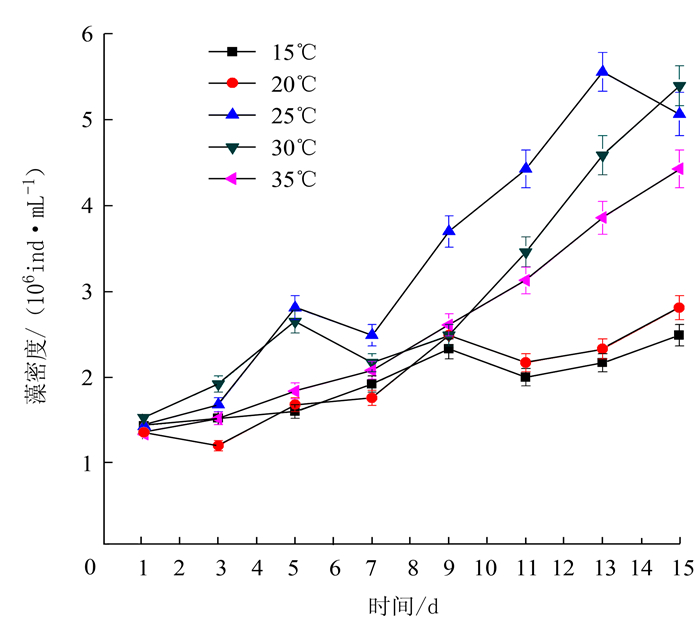
表 2 不同影响因素下藻最大比生长率及出现时间
Table 2 Maximum specific growth rate of chlorella and occurrence time under varied conditions
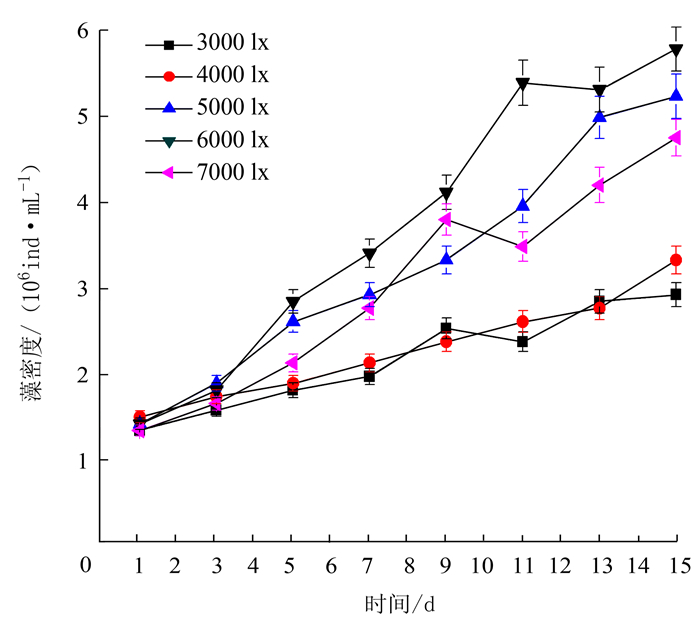
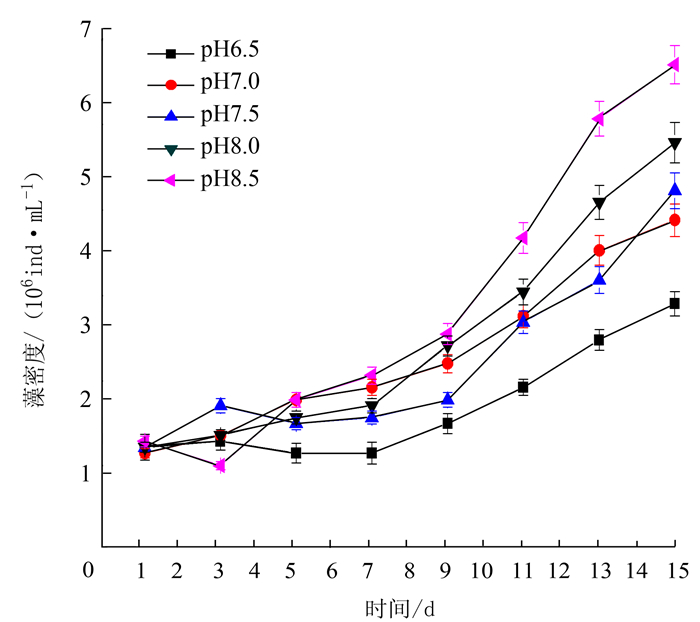
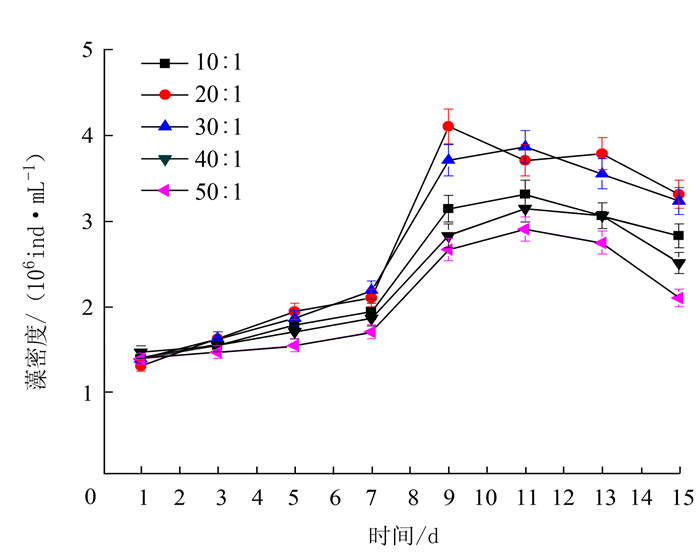
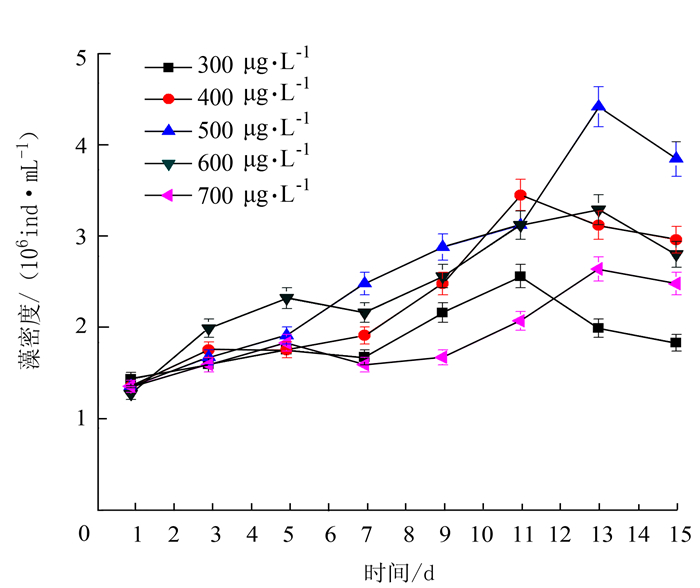
项目 处理 最大比生长率/% 出现时间/d 温度 15℃ 5.4 9~11 20℃ 6.8 9~11 25℃ 10.5 9~11 30℃ 11.2 9~11 35℃ 7.3 9~11 光照强度 3000 lx 5.6 9~11 4000 lx 7.4 9~11 5000 lx 10.0 9~11 6000 lx 12.6 9~11 7000 lx 12.1 9~11 pH 6.5 5.6 13~15 7.0 8.8 13~15 7.5 8.5 13~15 8.0 9.5 13~15 8.5 10.8 13~15 N/P 10:01 9.3 9~11 20:01 13.0 9~11 30:01 11.2 9~11 40:01 7.5 9~11 50:01 5.1 9~11 铁离子质量浓度 300 μg·L-1 5.3 9~11 400 μg·L-1 8.5 11~13 500 μg·L-1 9.1 13~15 600 μg·L-1 8.2 11~13 700 μg·L-1 5.2 13~15
下载: 导出CSV
表 3 正交试验结果
Table 3 Results of orthogonal experiment
因素 温度 光照 pH N/P 铁离子质量浓度 空列 藻密度 处理1 1 1 1 1 1 1 3.04 处理2 1 2 2 2 2 2 3.15 处理3 1 3 3 3 3 3 3.35 处理4 1 4 4 4 4 4 3.56 处理5 1 5 5 5 5 5 3.45 处理6 2 1 2 3 4 5 3.35 处理7 2 2 3 4 5 1 3.21 处理8 2 3 4 5 1 2 3.93 处理9 2 4 5 1 2 3 4.25 处理10 2 5 1 2 3 4 4.30 处理11 3 1 3 5 2 4 5.30 处理12 3 2 4 1 3 5 5.58 处理13 3 3 5 2 4 1 5.47 处理14 3 4 1 3 5 2 5.35 处理15 3 5 2 4 1 3 5.72 处理16 4 1 4 2 5 3 4.89 处理17 4 2 5 3 1 4 5.02 处理18 4 3 1 4 2 5 4.65 处理19 4 4 2 5 3 1 5.41 处理20 4 5 3 1 4 3 5.30 处理21 5 1 5 4 3 2 4.32 处理22 5 2 1 5 4 3 4.20 处理23 5 3 2 1 5 4 4.42 处理24 5 4 3 2 1 5 4.75 处理25 5 5 4 3 2 1 4.55 K1 3.310 4.194 4.330 4.530 4.490 4.348 K2 3.818 4.256 4.414 4.516 4.392 4.190 K3 5.494 4.364 4.412 4.324 4.604 4.642 K4 5.072 4.670 4.502 4.298 4.406 4.536 K5 4.468 4.678 4.504 4.494 4.270 4.356 极差 2.184 0.484 0.174 0.232 0.334 0.452
下载: 导出CSV
-
[1] 虞为, 李卓佳, 朱长波, 等.我国对虾生态养殖的发展现状、存在问题与对策[J].广东农业科学, 2011, 38(17):168-171. DOI: 10.3969/j.issn.1004-874X.2011.17.060 [2] 刘孝竹, 李卓佳, 曹煜成, 等.珠江三角洲低盐度虾池秋冬季浮游微藻群落结构特征的研究[J].农业环境科学学报, 2009, 28(5):1010-1018. https://www.wenkuxiazai.com/word/4f37eb5b10661ed9ad51f382-1.doc [3] 常布辉, 李兰, 姚磊.藻类培养与利用研究进展[J].水资源研究, 2013, 2(1):76-83. http://zhifu.cnki.net/Wechat.aspx [4] 曹煜成, 李卓佳, 梁伟峰, 等. 一种对虾养殖池藻相的构建方法: 中国, CN 101363004 B[P]. 2009-02-11. [5] 王起华, 施定基.几种海洋蓝藻生长条件的研究[J].海洋科学, 1997, 21(4):60-63. https://www.researchgate.net/profile/Jianming_Deng4/publication/286886727_Effects_of_enhanced_water_temperature_and_nutrient_concentration_on_algal_growth_in_winter_and_spring_season_in_Lake_Taihu_China/links/56a63e8f08aeca0fddcb4b57.pdf [6] 钱芸, 戴树桂, 刘广良, 等.硝酸镧对铜绿微囊藻生长特性的影响[J].中国环境科学, 2003, 23(1):7-11. http://www.cnki.com.cn/Article/CJFDTOTAL-ZGHJ200301002.htm [7] 贾含帅, 刘汉湖, 胡舒, 等. 4种环境因子对景观水体绿藻生长繁殖的影响[J].中国城市林业, 2011, 9(6):52-54. http://www.cqvip.com/qk/87510X/201106 [8] 蔡清海. 福建诏安湾环境质量评价[C]//全国海水养殖学术研讨会, 2007. [9] 江灵芝, 孙雪, 王玮蔚, 等.盐度对蛋白核小球藻生长、叶绿素荧光参数及代谢酶的影响[J].宁波大学学报(理工版), 2013(3):6-10. http://www.cqvip.com/QK/91297X/201303/46436559.html [10] 韩志国. 环境胁迫(盐胁, 热胁, 渗透胁迫)对两种海洋浮游植物的影响[D]. 广州: 暨南大学, 2002. http://cdmd.cnki.com.cn/Article/CDMD-10559-2002113814.htm [11] 郭亚丽, 傅海燕, 黄国和, 等.阿魏酸和香豆素对铜绿微囊藻的化感作用[J].环境科学, 2013, 34(4):1492-1497. http://industry.wanfangdata.com.cn/yj/Detail/Periodical?id=Periodical_hjkx201304040 [12] 董正臻, 董振芳, 丁德文.快速测定藻类生物量的方法探讨[J].海洋科学, 2004, 28(11):1-2. DOI: 10.3969/j.issn.1000-3096.2004.11.001 [13] 晁建颖, 颜润润, 张毅敏.不同温度下铜绿微囊藻和斜生栅藻的最佳生长率及竞争作用[J].生态与农村环境学, 2011, 27(2):53-57. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=ncsthj201102010 [14] 欧阳峥嵘, 温小斌, 耿亚红, 等.光照强度、温度、pH、盐度对小球藻(Chlorella)光合作用的影响[J].武汉植物学研究, 2010, 28(1):49-55. http://www.cnki.com.cn/Article/CJFDTOTAL-WZXY201001011.htm [15] WANG Z H, YUAN M L, LIANG Y, et al. Effects of temperature and organic and inorganic nutrients on the growth of Chattonella marina(Raphidophyceae) from the Daya Bay, South China Sea[J]. Acta Oceanologica Inica, 2011, 30(3):124-131. DOI: 10.1007/s13131-011-0127-2
[16] MARCIN W W, MARCIA K, JAMES J E, et al. Joint effect of phosphorus limitation and temperature on alkaline phosphatase activity and somatic growth in Daphnia magna[J].Oecologia, 2011, 165:837-846. DOI: 10.1007/s00442-010-1863-2
[17] 刘春光, 金相灿, 邱金泉, 等.光照与磷的交互作用对两种淡水藻类生长的影响[J].中国环境科学, 2005, 25(1):32-36. http://www.cnki.com.cn/Article/CJFDTOTAL-XCNY200302022.htm [18] 徐慧, 纪道斌, 崔玉洁, 等.不同光照强度对小球藻生长的影响[J].微生物学通报, 2016, 43(5):1027-1034. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=tjnykx201504025 [19] 刘春光, 金相灿, 孙凌, 等.水体pH和曝气方式对藻类生长的影响[J].环境污染与防治, 2006, 28(3):161-163. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=hjwryfz200603001 [20] 薛凌展, 黄种持, 林泽, 等.铜绿微囊藻和普通小球藻在不同pH下生长特性及竞争参数计算[J].福建农业学报, 2010, 25(2):142-148. http://www.cnki.com.cn/Article/CJFDTotal-SICA195903031.htm [21] RUANGSOMBOON S, GANMANEE M, CHOOCHOTE S. Effects of different nitrogen, phosphorus, and iron concentrations and salinity on lipid production in newly isolated strain of the tropical green microalga, Scenedesmus dimorphus KMITL.[J]. Journal of Applied Phycology, 2013, 25(3):867-874. DOI: 10.1007/s10811-012-9956-4
[22] 孟顺龙, 王菁, 裘丽萍, 等.氮磷质量浓度对普通小球藻和鱼腥藻生长竞争的影响[J].生态环境学报, 2015(4):658-664. http://www.cqvip.com/QK/97636C/201504/665024424.html [23] 曹煜成, 李卓佳, 胡晓娟, 等.磷浓度与氮磷比对蛋白核小球藻氮磷吸收效应的影响[J].生态科学, 2017, 36(5):34-40. https://wuxizazhi.cnki.net/qikan-WSWX200505017.html [24] 张晶晶, 周进, 张怀瑾, 等.不同营养条件下铜绿微囊藻(Microcystis aeruginosa)和小球藻(Chlorella vulgaris)的生长竞争行为[J].海洋与湖沼, 2016, 47(5):1013-1023. DOI: 10.11693/hyhz20160400085 [25] 郑晓宇, 顾詠洁, 金妍.不同氮、磷质量浓度下四尾栅藻的生长研究[J].生态环境学报, 2010, 19(11):2663-2668. DOI: 10.3969/j.issn.1674-5906.2010.11.025 [26] 董娟, 李大平, 陶勇, 等.氮磷胁迫下藻-菌群落的变化研究[J].环境科学与技术, 2012, 35(12):40-45, 55. DOI: 10.3969/j.issn.1003-6504.2012.12.009 [27] 李东侠, 丛威, 蔡昭铃, 等. Fe3+对赤潮异弯藻生长和光合作用的影响[J].应用与环境生物学报, 2003, 9(3):254-258. https://www.wenkuxiazai.com/word/00576a9fd5bbfd0a795673eb-1.doc [28] 迟光宇, 陈欣, 史奕, 等.铁对藻类生长的影响及其光谱识别研究展望[J].光谱学与光谱分析, 2009, 29(12):3344-3347. DOI: 10.3964/j.issn.1000-0593(2009)12-3344-04




计量
- 文章访问数: 1482
- HTML全文浏览量: 331
- PDF下载量: 18
